- ╩ūĒō Į±╚šäė(d©░ng)æB(t©żi) ╚╦▓┼╩ął÷ ą┬╝╝ąg(sh©┤)īŻÖ┌ ųąć°┐ŲīW(xu©”)╚╦ įŲš╣┼_(t©ói) įŲųv╠├ų▒▓ź Ģ■(hu©¼)š╣ųąą─ ╠žār(ji©ż)īŻÖ┌ ╝╝ąg(sh©┤)┐ņėŹ ├Ō┘M(f©©i)įćė├

-

╔·╬’═©╣┘╬ó
┼Ń─ŃūźūĪ╔·├³┐Ų╝╝
╠°äė(d©░ng)Ą─├}▓½
╔·╬’═©╣┘╬ó
┼Ń─ŃūźūĪ╔·├³┐Ų╝╝
╠°äė(d©░ng)Ą─├}▓½
RNAą▐’Śļ[▓žĄ─╔±├žš{(di©żo)┐ž
ĪŠūų¾wŻ║ ┤¾ ųą ąĪ Ī┐ Ģr(sh©¬)ķgŻ║2021─Ļ07į┬09╚š üĒį┤Ż║╔·╬’═©
ŠÄ▌ŗ═Ų╦]Ż║
ĪĪĪĪ▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)įĮüĒįĮ╩▄ĄĮ┐ŲīW(xu©”)╝ęéāĄ─ųžęĢŻ¼│╔×ķ┴╦Į³üĒ┼dŲĄ─¤ßķTŅI(l©½ng)ė“ų«ę╗ĪŻŲ∙Į±×ķų╣Ż¼į┌RNA╔Žęč░l(f©Ī)¼F(xi©żn)┴╦170ČÓĘN╗»īW(xu©”)ą▐’Ś[1]ĪŻ
2016─ĻĄ─ūŅ║¾ę╗╠ņŻ¼Nature Methods┌sų°░l(f©Ī)▓╝┴╦2016─ĻČ╚╝╝ąg(sh©┤)Ż¼▓╗╔┘╚╦šJ(r©©n)×ķ«ö(d©Īng)─ĻĄ──ĻČ╚╝╝ąg(sh©┤)ę╗Č©╩Ū┤¾╗Ą─CRISPR╝╝ąg(sh©┤)Ż¼╚╗▓ó┬čŻ¼ī”(du©¼)ė┌ŅHŠ▀Ū░š░ąįĄ──ĻČ╚╝╝ąg(sh©┤)▒P³c(di©Żn)üĒšfŻ¼ę╗éĆ(g©©)▓ó▓╗│ŻęŖĄ─├¹į~Ż║Epitranscriptome analysisŻ©▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)Ż®▓┼╩Ūš²ĮŌĪŻ
Epitranscriptome analysis▀@éĆ(g©©)├¹ĘQ╩Ūė╔ŽŻ┼DšZĪ░epiĪ▒ū„×ķŪ░ŠYŻ¼ųĖĄ─Š═╩Ū│²ķ_ęčų¬╣”─▄╗“▀zé„ąįŻ¼╚╬║╬╠Ē╝ėĄĮ║╦▄š╦ß╔ŽĄ─ą▐’ŚĪŻÄū╩«─ĻüĒŻ¼┐ŲīW(xu©”)╝ęéāÄū║§Č╝ø]ėąūóęŌĄĮRNAą▐’ŚŻ¼ę“?y©żn)ķįńį┌╔ŽéĆ(g©©)╩└╝o(j©¼)60─Ļ┤·║═70─Ļ┤·RNA╔ŽĄ─ś╦(bi©Īo)ėøŠ═▒╗░l(f©Ī)¼F(xi©żn)┴╦Ż¼Ą½╩Ū┤¾╝ęų╗ĻP(gu©Īn)ūóė┌tRNA║═rRNAŻ¼ęį╝░DNA╔ŽĄ─▒Ēė^▀zé„ą▐’ŚĪŻ
Ą½ļSų°┐ŲīW(xu©”)╝ęéā░l(f©Ī)¼F(xi©żn)┴╦│÷¼F(xi©żn)į┌╦∙ėąRNAĘNŅÉųąĄ─╗»īW(xu©”)ś╦(bi©Īo)ėøŻ¼äė(d©░ng)æB(t©żi)╠Ē╝ė╗“š▀╚ź│²▀@ą®ś╦(bi©Īo)ėøĄ─Ī░Readerīæ╩ųĪ▒║═Ī░EraserŽŲż▓┴Ī▒Ż¼ųžą┬³c(di©Żn)╚╝┴╦ī”(du©¼)RNAą▐’ŚĄ─┼d╚żĪŻ└²╚ńŻ¼Å─Ž┘Óč▀╩╔Ž╚ź│²ę╗éĆ(g©©)╝ū╗∙╗∙łF(tu©ón)Ą─├ĖŻ¼┼c░óĀ¢┤─║Ż─¼░Y╗╝▓Ī’L(f©źng)ļU(xi©Żn)ų«ķgĄ─ĻP(gu©Īn)┬ō(li©ón)Ż¼▒Ē├„┴╦▀@ĘNą▐’Śį┌╔±Įø(j©®ng)ĮĪ┐ĄĘĮ├µ░ńč▌┴╦ųžę¬š{(di©żo)╣Ø(ji©”)ū„ė├ĪŻ
ė╔┤╦▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)įĮüĒįĮ╩▄ĄĮ┐ŲīW(xu©”)╝ęéāĄ─ųžęĢŻ¼│╔×ķ┴╦Į³üĒ┼dŲĄ─¤ßķTŅI(l©½ng)ė“ų«ę╗ĪŻŲ∙Į±×ķų╣Ż¼į┌RNA╔Žęč░l(f©Ī)¼F(xi©żn)┴╦170ČÓĘN╗»īW(xu©”)ą▐’Ś[1]ĪŻ▀@ą®ą▐’Ś┤¾┴┐Ęų▓╝į┌ĘŪŠÄ┤aRNAŻ©ncRNAŻ®Ż¼╠žäe╩ŪrRNA, tRNA║═snRNA╔ŽŻ¼×ķncRNAį┌ĘŁūg┼c╝¶Įėųą░l(f©Ī)ō]š²│Ż╣”─▄╦∙▒žąĶĪŻ┴Ņ╚╦┼dŖ^Ą─╩ŪŻ¼čąŠ┐╚╦åT░l(f©Ī)¼F(xi©żn)m6AŻ©N6-methyladenosineŻ®Ż¼m1AŻ©N1-methyladenosineŻ®Ż¼m5CŻ©5-methylcytidineŻ®Ż¼hm5CŻ©5-hydroxylmethylcytidineŻ®Ż¼IŻ©inosineŻ®ęį╝░”ūŻ©pseudouridineŻ®Ą╚╗»īW(xu©”)ą▐’Śę▓Ęų▓╝į┌šµ║╦╔·╬’mRNA╔ŽŻ¼ė░ĒæmRNAĄ─┤·ųx┼c╣”─▄ĪŻ╠žäe╩Ū░ķļSų°įSČÓmRNAą▐’Ś├ĖŻ©WriterŻ®Īó╚źą▐’Ś├ĖŻ©EraserŻ®║═ą▐’ŚūR(sh©¬)äeĄ░░ūŻ©ReaderŻ®Ą─ą┬░l(f©Ī)¼F(xi©żn)Ż¼mRNA╗»īW(xu©”)ą▐’ŚĄ─┐╔─µūā╗»┼cäė(d©░ng)æB(t©żi)š{(di©żo)┐žųžą┬╝żŲ┴╦蹊┐╚╦åTĄ─┼d╚żĪŻ
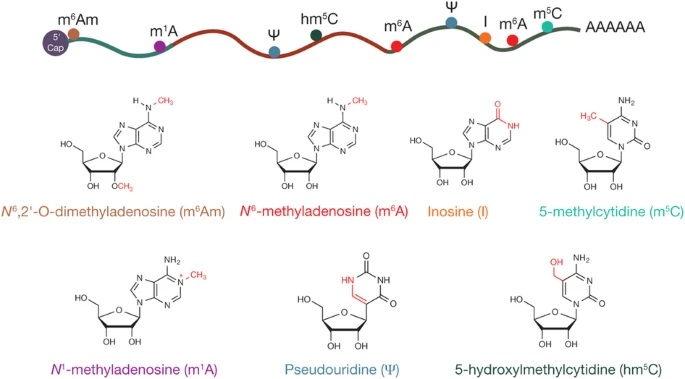
łDę╗üĒūį[2]
▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)蹊┐ĘČć·
▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)Ż©epitranscriptomicsŻ¼ėųĘQĪ░RNA▒Ēė^▀zé„īW(xu©”)Ī▒Ż®╩ŪųĖ▐D(zhu©Żn)õø║¾ RNA ą▐’ŚŻ¼▀@ą®ą▐’ŚĮo▐D(zhu©Żn)õøĮMĦüĒ┴╦╣”─▄ŽÓĻP(gu©Īn)Ą─ūā╗»ĪŻ▒Ēė^▐D(zhu©Żn)õøĮMą▐’Ś░³└©ÄūéĆ(g©©)ųžę¬Ą─ RNA ╝ė╣ż╩┬╝■Ż¼░³└© RNA ŠÄ▌ŗĪó╝ū╗∙╗»║═╝¶ĮėŻ©łDČ■)ĪŻ

łDČ■üĒūį[4]
Å─╩ŪʱŠÄ┤aĄ░░ū┘|(zh©¼)üĒšfŻ¼RNA┐╔ęįĘų×ķŠÄ┤aRNAŻ©coding RNAŻ®ą▐’Ś║═ĘŪŠÄ┤aRNAŻ©non-coding RNA, ncRNAŻ®ā╔┤¾ŅÉĪŻŪ░š▀Š═╩ŪųĖmRNAŻ¼║¾š▀ät░³└©║▄ČÓĘNŅÉŻ¼╚ń▒Ŗ╦∙ų▄ų¬Ą─tRNA║═rRNAŻ¼ģó┼cRNAą▐’ŚĄ─snoRNAĄ╚ĪŻ
mRNAą▐’ŚčąŠ┐ūŅČÓĄ─Š═╩Ūm6A ą▐’ŚŻ¼įńį┌ 20 ╩└╝o(j©¼) 70 ─Ļ┤·Ż¼┐ŲīW(xu©”)╝ęéāŠ═į┌ RNA ųą░l(f©Ī)¼F(xi©żn)┴╦ m6A ą▐’ŚŻ¼ļS║¾įĮüĒįĮČÓĄ─蹊┐ūC├„m6A ą▐’ŚĄ─ųžę¬ąįŻ║m6A ą▐’Ś║═ mRNA Ą─ĘĆ(w©¦n)Č©ąįĪó╝¶Įė╝ė╣żĪóĘŁūgęį╝░ microRNA Ą─╝ė╣żėąĻP(gu©Īn)Ż╗m6A ▀Ć║═Ė╔╝Ü(x©¼)░¹├³▀\(y©┤n)Īó╔·╬’╣Ø(ji©”)┬╔ŽÓĻP(gu©Īn)Ż¼┐╔ęį┤┘╩╣Ė╔╝Ü(x©¼)░¹Å─ūį╬ęĖ³ą┬ĀŅæB(t©żi)▐D(zhu©Żn)Ž“╝Ü(x©¼)░¹Ęų╗»Ż¼čąŠ┐╚╦åT░l(f©Ī)¼F(xi©żn)Ż¼╝ū╗∙╗»Ģ■(hu©¼)┐sČ╠ mRNA Ą─░ļ╦źŲ┌Ż¼£p╔┘ŲõžSČ╚ĪŻ┐╔ęįšfŻ¼m6A ą▐’ŚÄū║§ė░Ēæ RNA ┤·ųxĄ─├┐éĆ(g©©)▓Į¾EĪŻ
Č°Į³─ĻüĒŻ¼ļSų°čąŠ┐Ą─╔Ņ╚ļŻ¼▓╗╔┘蹊┐╣żū„ę▓Å─mRNA▐D(zhu©Żn)×ķĻP(gu©Īn)ūóĘŪŠÄ┤aRNAĄ─╝ū╗∙╗»ī”(du©¼)ė┌╝▓▓Ī░l(f©Ī)╔·░l(f©Ī)š╣▀^│╠Ą─ųžę¬ū„ė├Ż¼░l(f©Ī)¼F(xi©żn)ĘŪŠÄ┤aRNAĄ─m6A╝ū╗∙╗»Ģ■(hu©¼)į┌Ė╔╝Ü(x©¼)░¹Ęų╗»Īó░®╝Ü(x©¼)░¹į÷ų│Ą╚▀^│╠ųąŲĻP(gu©Īn)µIū„ė├ĪŻ═¼mRNAę╗śėŻ¼lncRNA╔Žę▓┤µį┌ų°ČÓĘN╗»īW(xu©”)ą▐’ŚŻ¼m6A╝ū╗∙╗»ī”(du©¼)ė┌lncRNAüĒųvŻ¼┐╔ęįš{(di©żo)┐žlncRNAČ■╝ē(j©¬)ĮY(ji©”)śŗ(g©░u)Ż¼lncRNAĮY(ji©”)║ŽĄ░░ūŻ¼ęį╝░lncRNAĄ─ceRNAÖC(j©®)ųŲŻ¼░ą╗∙ę“Ą─m6Aą▐’ŚĪŻ
┤╦═Ō▀ĆėąŁh(hu©ón)ĀŅRNA╔Žm6Aą▐’ŚŻ¼▀@─▄ė░ĒæcircRNA║═RNAĮY(ji©”)║ŽĄ░░ū(RBP)ų«ķgĄ─ŽÓ╗źū„ė├Ż¼ęį╝░ś╦(bi©Īo)ėøā╚(n©©i)į┤RNAŻ¼Å─Č°īóŲõ┼c═Ōį┤RNAģ^(q©▒)Ęųķ_Ż¼▒▄├Ō▒╗ūį╔Ē├Ōę▀ŽĄĮy(t©»ng)ūR(sh©¬)äe╣źō¶ĪŻ
Small RNAŻ¼└²╚ńmicroRNAŻ¼tRNAüĒį┤ąĪRNA(tsRNAŻ¼░³└©tRF&tiRNA)Ą╚Ż¼ŲõRNAĘųūė╔ŽŠ▀ėąČÓĘN▓╗═¼Ą─ą▐’ŚŻ¼▀@ą®ą▐’Śę╗ĘĮ├µ─▄ē“š{(di©żo)┐žsmall RNAĄ─╗ŅąįŻ¼┴Ēę╗ĘĮ├µę▓┐╔ęį┘xėĶ╦³éāą┬Ą─╣”─▄ĪŻęčų¬▀@ą®RNAą▐’Ś═©▀^ČÓĘNĘųūėÖC(j©®)ųŲüĒ░l(f©Ī)ō]╣”─▄Ż¼╚ńRNAą▐’Ś┐╔ęįĖ─ūāmiRNAĄ─░ąŽ“ąį╗“Ė─ūātsRNA(tRF&tiRNA)┼cRNAĮY(ji©”)║ŽĄ░░ūĄ─ėH║═┴”Ż¼▀M(j©¼n)Č°░l(f©Ī)ō]╔·╬’╗ŅąįĄ╚ĪŻSmall RNAą▐’ŚūVĘų╬÷╩Ū▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)蹊┐Ą─ą┬Ū░螯¼Š▀ėąųžę¬Ą─┐ŲīW(xu©”)ęŌ┴x║═┼R┤▓ār(ji©ż)ųĄĪŻ
RNA ą▐’ŚŅI(l©½ng)ė“ūŅųžę¬Ą─│╔Š═
ī”(du©¼)ė┌Ų∙Į±×ķų╣RNAą▐’ŚŅI(l©½ng)ė“ūŅųžę¬Ą─│╔Š═Ż¼┐Ą─╬Ā¢┤¾īW(xu©”)Samie R. JaffreyĮ╠╩┌šJ(r©©n)×ķŻ¼Ī░Ą┌ę╗éĆ(g©©)ė├ė┌└LųŲ m6A╚½▐D(zhu©Żn)õøĮMĘĮĘ©┐╔─▄╩Ū▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)ŅI(l©½ng)ė“ųąūŅųžę¬Ą─╩┬╝■Ż¼¼F(xi©żn)į┌ęčĮø(j©®ng)▒╗Å═(f©┤)ųŲė├ė┌Ųõ╦¹ÄūĘN║╦▄š╦ßą▐’ŚĪŻ į┌╚½▐D(zhu©Żn)õøĮMū„łDų«Ū░Ż¼═©▀^╩╣ė├┘|(zh©¼)ūVĘ©╗“Ųõ╦¹Ęų╬÷£y┴┐Öz£y╦«ĮŌ RNA ųąą▐’ŚĄ─║╦▄š╦߯¼░l(f©Ī)¼F(xi©żn)ą▐’ŚĄ─║╦▄š╦ßĪŻ▀@ą®ĘĮĘ©╩Ū─Ż└Ōā╔┐╔Ą─Ż¼ė╚Ųõ╩Ūī”(du©¼)ė┌Ą═žSČ╚Ą─ą▐’ŚŻ║╝┤╩╣─Ńėąę╗éĆ(g©©)Ė▀╝āČ╚Ą─ mRNA ųŲ䮯¼─Ń▀Ć╩Ūō·(d©Īn)ą─╬ó┴┐Ą─╬█╚Š▐D(zhu©Żn)ęŲ RNA (tRNA) ╗“║╦╠Ū¾w RNA (rRNA) ┐╔─▄╩Ūą▐’ŚĄ─üĒį┤ĪŻ▀@╩╣Ą├║▄ļy┤_Č©ą▐’ŚĄ─║╦▄š╦ß╩ŪʱüĒūį mRNAĪŻĪ▒
ęį╔½┴ą╠ž└ŁŠSĘ“┤¾īW(xu©”)ßt(y©®)īW(xu©”)į║Ą─Gideon RechaviĮ╠╩┌ät▒Ē╩ŠŻ¼Ī░į┌╬ę┐┤üĒŻ¼įōŅI(l©½ng)ė“Ą─ų„ę¬│╔Š═╩Ū═©▀^mRNA ą▐’ŚĮę╩Š┴╦ę╗éĆ(g©©)ą┬Ą─ĪóÅ═(f©┤)ļsĄ─ĪóĖ▀Č╚├¶ĖąĄ─Īó┐╔š{(di©żo)╣Ø(ji©”)Ą─╗∙ę“▒Ē▀_(d©ó)š{(di©żo)┐žīėĪŻ▀@ę╗ą┬Ą─š{(di©żo)┐žīė└¹ė├┴╦mRNA Ą─¬Ü(d©▓)╠ž╠žąįĪ¬Ī¬╝┤╦³╩ŪČ╠Ģ║Ą─ĪóĖ▀Č╚ĮY(ji©”)śŗ(g©░u)╗»Ą─Īóį┌╝Ü(x©¼)░¹ģ^(q©▒)╩ęų«ķgęŲäė(d©░ng)▓ó═©▀^▐D(zhu©Żn)õøĘ┼┤¾Ą─ĪŻ▀@ą®ė░Ēæ▓┐Ęų╩Ūė╔Ī░ķåūxŲ„Ī▒Įķī¦(d©Żo)Ą─Ż¼└²╚ń╝ū╗∙╠ž«ÉąįĮY(ji©”)║ŽĄ░░ūŻ¼╦³Ą─ĶbČ©╩Ūą▐’ŚŅI(l©½ng)ė“Ą─ę╗éĆ(g©©)└’│╠▒«ĪŻ╗∙ę“▒Ē▀_(d©ó)Ą─š{(di©żo)╣Ø(ji©”)ę▓═©▀^Ī░īæ╩ųĪ▒║═Ī░ŽŲż▓┴Ī▒Ą─ą▐Ė─░▓čb║═äh│²ų«ķgĄ─ŽÓ╗źū„ė├▀M(j©¼n)ąąš{(di©żo)š¹ĪŻį┌▀^╚źĄ─╩«─ĻųąŻ¼│÷¼F(xi©żn)┴╦ÄūéĆ(g©©)ųžę¬Ą─Į╠ė¢(x©┤n)ĪŻ╩ūŽ╚Ż¼mRNA ą▐’ŚĘŪ│ŻŲš▒ķŻ¼öĄ(sh©┤)Ū¦éĆ(g©©)╗∙ę“▐D(zhu©Żn)õø▒Š▒╗ą▐’ŚĪŻėą╚żĄ─╩ŪŻ¼ę╗ą®ą▐Ė─Š█╝»į┌╠žČ©Ą─▐D(zhu©Żn)õø╬╗ų├Ż╗└²╚ńŻ¼╝Ī▄šų„ę¬┤µį┌ė┌ųžÅ═(f©┤)Ą─ Alu ą“┴ąųąŻ¼m6A ā×(y©Łu)Ž╚ą▐’ŚĮKų╣├▄┤aūėĖĮĮ³║═ā╚(n©©i)▓┐═Ō’@ūėĪó AUG Ų╩╝├▄┤aūėų▄ć·Ą─ m1A ┤žŻ¼▀@▒Ē├„├┐éĆ(g©©)ą▐’Ś═©▀^▓╗═¼Ą──Ż╩ĮŲū„ė├ąąäė(d©░ng)ĪŻ┤╦═ŌŻ¼ę╗ą®ą▐’ŚŻ¼╚ń m6A ║═ m1AŻ¼į┌╚╦║═ąĪ╩¾ų«ķg▒Ē¼F(xi©żn)│÷Ė▀Č╚Ą─▒Ż╩žąįĪŻ
┴Ēę╗éĆ(g©©)ųžę¬Ą─│╔Š═╩Ū░l(f©Ī)¼F(xi©żn)╠žČ©Ą─ą▐Ė─┐╔ęį═©▀^▓╗═¼Ą─ąą×ķ─Ż╩ĮŻ¼═©▀^▓╗═¼Ą─ūx╚ĪŻ¼ę└┘ćė┌╔ŽŽ┬ė╬ĪŻ▀Ćėąę╗éĆ(g©©)ųžę¬░l(f©Ī)¼F(xi©żn)╩Ūę╗ą® mRNA ą▐’ŚĄ─äė(d©░ng)æB(t©żi)╠žąįŻ¼┐╔ęįī”(du©¼)Łh(hu©ón)Š│┤╠╝żū÷│÷┐ņ╦┘Ę┤æ¬(y©®ng)Ż╗ m6A ║═ m1A ęčĮø(j©®ng)ūC├„┴╦▀@ĘNäė(d©░ng)æB(t©żi)╠žąįĪŻ mRNA ą▐’ŚĄ─║╦ą─ū„ė├¾w¼F(xi©żn)į┌«É│Żą▐’Śī”(du©¼)╚╦ŅÉ║═ąĪ╩¾įńŲ┌░l(f©Ī)ė²ęį╝░╚╦ŅÉ░®░YĪóčū░Y║═╔±Įø(j©®ng)ūāąįĄ─ŲŲē─ąįė░ĒæŻ¼▀M(j©¼n)ę╗▓ĮÅŖ(qi©óng)š{(di©żo)┴╦▀@ę╗š{(di©żo)╣Ø(ji©”)īėĄ─ųžę¬ąįĪŻĪ▒
¢|Š®┤¾īW(xu©”)Ą─Tsutomu SuzukiĮ╠╩┌▒Ē╩ŠŻ¼Ī░ęįŪ░ĻP(gu©Īn)ė┌ RNA ą▐’ŚĄ─╔·╗»čąŠ┐ų„ę¬╝»ųąį┌Įø(j©®ng)ĄõĄ─ĘŪŠÄ┤a RNAŻ¼░³└© tRNAĪórRNA ║═ąĪ║╦ RNA (snRNA)Ż¼ę“?y©żn)ķ▀@ą®RNA į┌╝Ü(x©¼)░¹ųą║¼┴┐žSĖ╗ĪŻ╚╗Č°Ż¼ūŅĮ³Ż¼╩╣ė├ NGS ╝╝ąg(sh©┤)ī”(du©¼) RNAą▐’Ś▀M(j©¼n)ąą╚½▐D(zhu©Żn)õøĮMĘų╬÷ęčČ©│÷ÄūĘNēA╗∙ą▐’ŚŻ¼░³└© mRNA ųąĄ─╝Ī▄š (I)Īóm6AĪóm5CĪó”Ę ║═ m1A ęį╝░ķLµ£ĘŪŠÄ┤a RNAĪŻ▀@Ę┤▀^üĒ▀@┤¾┤¾═žīÆ┴╦▒Ēė^▐D(zhu©Żn)õøĮMĄ─Ė┼─ŅĪŻ
į┌▀^╚źĄ─╩«Äū─ĻųąŻ¼═©▀^Ę┤Ž“▀zé„īW(xu©”)║═┘|(zh©¼)ūVĘ©Ż¼┐ŲīW(xu©”)╝ęéāūR(sh©¬)äe│÷┴╦┬±▓žį┌─Ż╩Į╔·╬’╗∙ę“ĮMųąĄ─RNA ą▐’Ś├ĖĪŻ╩╣ė├▀@ĘNĘĮĘ©Ż¼╬ęéā│╔╣”ĶbČ©┴╦ 40 éĆ(g©©)RNA ą▐’Ś╗∙ę“ĪŻ
▀ĆųĄĄ├ę╗╠ߥ─╩ŪŻ¼┼c╝▓▓ĪŽÓĻP(gu©Īn)Ą─═Ō’@ūėĮM£yą“ėąų·ė┌ĮŌ╬÷RNA ą▐’Ś├Ė═╗ūā?n©©i)ń║╬ę²░l(f©Ī)┴╦įSČÓ╚╦ŅÉ╝▓▓ĪĪŻĪ▒[5]
Å──┐Ū░Ą─蹊┐│╔╣¹üĒ┐┤Ż¼▒Ēė^▐D(zhu©Żn)õøĮMė░Ēæ╔Ņ▀h(yu©Żn)Ż¼╩Ūū„×ķīó┐╔╦▄ąį»B╝ėį┌Ųõ╦¹╗∙ę“ĮM╔Ž▀BŠĆ▐D(zhu©Żn)õøĮM╔ŽĄ─ę╗ĘN╩ųČ╬ĪŻ▒Ēė^▐D(zhu©Żn)õøĮMĪ░┤·┤aĪ▒▓╗āH┐╔ęįåóė├╗“į÷ÅŖ(qi©óng) RNA ┤▀╗»╗“ RNA ę└┘ćąįĘ┤æ¬(y©®ng)ųąĄ─╠žČ©╗»īW(xu©”)Ę┤æ¬(y©®ng)Ż¼▀Ć┐╔ęįĖ─ūā RNA ĮY(ji©”)śŗ(g©░u)-╣”─▄ĻP(gu©Īn)ŽĄŻ¼Å─Č°ęįĢr(sh©¬)┐š║═ą┼╠¢(h©żo)ę└┘ćąįĘĮ╩Į╠ß╣®Ņ~═ŌĄ─╗∙ę“š{(di©żo)┐žīėĪŻ
╚╗Č°Ż¼▒Ēė^▐D(zhu©Żn)õøĮMĄ─╣”─▄蹊┐┬õ║¾ė┌▒Ēė^╗∙ę“ĮMĄ─╣”─▄蹊┐Ż¼▀@╩Ūę“?y©żn)ķ╚▒Ę”┐╔ęįį┌▐D(zhu©Żn)õøĮMĘČć·ā╚(n©©i)Öz£y▀@ą®▒Ēė^▐D(zhu©Żn)õøĮMś╦(bi©Īo)ėøĄ─ņ`├¶ŪęĘĆ(w©¦n)ĮĪĄ─╝╝ąg(sh©┤)ĪŻčąŠ┐▒Ēė^▐D(zhu©Żn)õøĮM┤µį┌ÄūéĆ(g©©)ų„ę¬╠¶æ(zh©żn)ĪŻ╩ūŽ╚Ż¼┤¾ČÓöĄ(sh©┤) RNA ą▐’Ś▓╗─▄═©▀^Ė▀═©┴┐£yą“ų▒ĮėÖz£yĄĮĪŻę“?y©żn)ķī?du©¼) RNA Ą─╗»īW(xu©”)ą▐’Ś═©│Ż▓╗Ģ■(hu©¼)Ė─ūāą▐’ŚēA╗∙Ą─ēA╗∙┼õī”(du©¼)╠žąįŻ¼─µ▐D(zhu©Żn)õø (RT) īó║åå╬Ąž▓┴│²▀@ą®ą▐’ŚŻ¼▓ó╩╣╦³éā┼c│ŻęÄ(gu©®) RNA ēA╗∙¤oĘ©ģ^(q©▒)ĘųĪŻŲõ┤╬Ż¼ļm╚╗ rRNAĪótRNA ║═ snRNA ║▄žSĖ╗Ż¼Ą½Ųõ╦¹ŅÉą═Ą─RNAŻ¼└²╚ń mRNA ║═ķLµ£ĘŪŠÄ┤a RNA (lncRNA)Ż¼žSČ╚┐╔─▄▌^Ą═ĪŻĄ┌╚²Ż¼╚▒Ę”¼F(xi©żn)ėąĄ─ėŗ(j©¼)╦Ń╣żŠ▀üĒ┤┘▀M(j©¼n)Å─£yą“öĄ(sh©┤)ō■(j©┤)ūR(sh©¬)äeą▐’Ś╬╗³c(di©Żn)Ą──▄┴”ĪŻ
ąę▀\(y©┤n)Ą─╩ŪŻ¼Į³─ĻüĒŻ¼ßśī”(du©¼)▓╗═¼▒Ēė^▐D(zhu©Żn)õøĮMĄ─蹊┐ĘĮĘ©╚ĪĄ├┴╦ųž┤¾▀M(j©¼n)š╣ĪŻ▀@ą®ą┬╣żŠ▀Ä═ų·čąŠ┐╚╦åT┤_Č© RNA ą▐’ŚĄ─╬╗ų├Ż¼▓óĮę╩Š▀@ą®ą▐’Śį┌š¹éĆ(g©©)▐D(zhu©Żn)õøĮMųąĄ─▓╗═¼Ęų▓╝─Ż╩ĮĪŻ«ö(d©Īng)▀@ą®ĘĮĘ©┼cŲõ╦¹ą┬┼d╣żŠ▀Ż©└²╚ń╗∙ę“ĮMŠÄ▌ŗ╣żŠ▀Ż®ĮY(ji©”)║Ž╩╣ė├Ģr(sh©¬)Ż¼RNA ą▐’Ś├ĖĄ─░ąś╦(bi©Īo)Š═ęč┤_Č©ĪŻ┤╦═ŌŻ¼▀@ą®╝╝ąg(sh©┤)▀ĆĮę╩Š┴╦▓╗═¼╔·└Ē?xi©żng)l╝■Ž┬▓╗═¼▒Ēė^▐D(zhu©Żn)õøĮMś╦(bi©Īo)ėøĄ─äė(d©░ng)æB(t©żi)ąį┘|(zh©¼)ĪŻ═¼Ģr(sh©¬)▀@ą®╣żŠ▀╩╣╚╦éā─▄ē“░l(f©Ī)¼F(xi©żn)▀xō±ąįūR(sh©¬)äe╠žČ©▒Ēė^▐D(zhu©Żn)õøĮMś╦(bi©Īo)ėø▓ó┤_Č©Ųõ╣”─▄Ą─Ī░ķåūxŲ„Ī▒Ą░░ūĪŻę“┤╦Ż¼ą┬Ą─蹊┐╣żŠ▀▓╗āH┐╔ęįī”(du©¼)▒Ēė^▐D(zhu©Żn)õøĮM▀M(j©¼n)ąą╚½├µĘų╬÷Ż¼Č°Ūę▀Ć╩Ū╣”─▄▒Ēė^▐D(zhu©Żn)õøĮM蹊┐Ą─īÜ┘F┘Yį┤ [2]ĪŻ
ó┼ Öz£yĘų╬÷ĘĮĘ©
蹊┐ RNA ą▐’ŚĄ─ĘĮĘ©░³└©╩Ūę║ŽÓ╔½ūV(LC-MS) Īó╗∙ė┌Ė▀═©┴┐£yą“Ą─ĘĮĘ©║═ąŠŲ¼Ęų╬÷Ą╚ĘĮĘ©ĪŻ
ó┘ mRNA&lncRNA▒Ēė^▐D(zhu©Żn)õøĮMąŠŲ¼
RNAą▐’ŚĄ─Øōį┌╣”─▄▓╗āH╚ĪøQė┌Ųõ╦∙ą▐’ŚĄ─╩Ū║╬ĘN╗∙ę“▐D(zhu©Żn)õø▒ŠŻ¼═¼Ģr(sh©¬)ę▓╚ĪøQė┌▒╗ą▐’Ś▓┐Ęųį┌įō▐D(zhu©Żn)õø▒Šųą╦∙š╝Ą─░┘Ęų▒╚ĪŻ╚╗Č°Ż¼─┐Ū░┤¾▓┐Ęų▐D(zhu©Żn)õøĮM╦«ŲĮĄ─Ą─RNAą▐’ŚÖz£yĘĮĘ©ų°ųžė┌īżšę▐D(zhu©Żn)õø▒Š╔ŽĄ─ą▐’Ś╬╗³c(di©Żn)Ż¼▓╗─▄ē“Č©┴┐ĄžÖz£y▒╗ą▐’Ś▐D(zhu©Żn)õø▒ŠĄ─░┘Ęų▒╚ĪŻ▀@ę╗ŅÉČ©┴┐ą┼ŽóĄ─╚▒Ę”ęčę²ŲįĮüĒįĮČÓ┐Ųčą╣żū„š▀Ą─ĻP(gu©Īn)ūóĪŻ
mRNAą▐’ŚĄ─Øōį┌ė░Ēæ╝╚╚ĪøQė┌ŲõĘųūėą¦æ¬(y©®ng)Ż¼ę▓╚ĪøQė┌▒╗ą▐’Ś▐D(zhu©Żn)õø▒ŠĄ─░┘Ęų▒╚ĪŻ└²╚ńŻ¼ę╗ĘN┐╔ęį╝ė╦┘mRNAĮĄĮŌĄ─ą▐’ŚŻ¼╚ń╣¹ų╗ėą1%Ą─▐D(zhu©Żn)õø▒Š▒╗ą▐’ŚŻ¼’@╚╗▓╗╠½┐╔─▄«a(ch©Żn)╔·╚╬║╬╔·╬’╣”─▄Ż¼╚╗Č°«ö(d©Īng)ę╗ĘNą▐’Ś┐╔ęį┤┘╩╣mRNAĘŁūg│╔ą┬Ą─Ą░░ūüåą═Ģr(sh©¬)Ż¼╝┤╩╣ą▐’Ś╦«ŲĮ║▄Ą═Ż¼ę▓┐╔─▄«a(ch©Żn)╔·ųžę¬Ą─╔·╬’╣”─▄ĪŻ«ö(d©Īng)Ū░Ą─m6A║═”ĘÖz£yĘĮĘ©ŠųŽ▐ąįį┌ė┌╚▒Ę”ą▐’Ś│╠Č╚Ą─Č©┴┐ą┼ŽóĪŻm6AĄ─š{(di©żo)┐žū„ė├┐╔ęį═©▀^pulldown m6AĄ─ĘĮĘ©Ż¼Öz£y╠žČ©ą“┴ąį┌▓╗═¼ĀŅæB(t©żi)Ž┬Ą─ŽÓī”(du©¼)Ė╗╝»│╠Č╚▀M(j©¼n)ąą═ŲöÓŻ¼Ą½▓╗─▄Å─▀@ą®öĄ(sh©┤)ō■(j©┤)ųą½@Ą├▒╗ą▐’ŚmRNAĄ─Į^ī”(du©¼)┴┐ĪŻą┬Ą─┐╔Č©┴┐m6A║═”ĘĄ─Ė▀═©┴┐Öz£yĘĮĘ©Ż¼īóśO┤¾Ą─┤┘▀M(j©¼n)įōŅI(l©½ng)ė“Ą─░l(f©Ī)š╣[7]ĪŻ
┴Ēę╗éĆ(g©©)ųžę¬Ą─å¢Ņ}╩ŪĻU├„RNAą▐’ŚĄ─╗»īW(xu©”)ėŗ(j©¼)┴┐Ą─äė(d©░ng)æB(t©żi)ūā╗»ĪŻ─┐Ū░Ż¼▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)蹊┐Ą─ųž³c(di©Żn)┤¾ČÓ╩Ū──ą®╬╗³c(di©Żn)▒╗ą▐’ŚŻ¼Č°▓╗╩ŪRNAųą├┐éĆ(g©©)▒╗ą▐’Ś╬╗³c(di©Żn)Ą─š╝▒╚ĪŻĄ══©┴┐Ęų╬÷mRNA║═▓ĪČŠRNAĄ─m6Aą▐’Ś╬╗³c(di©Żn)’@╩ŠŻ¼╚╬║╬m6Aą▐’Ś╬╗³c(di©Żn)Ą─š╝▒╚Č╝▓╗Ģ■(hu©¼)▀_(d©ó)ĄĮ100%ĪŻą▐’ŚĄ─╗»īW(xu©”)ėŗ(j©¼)┴┐ūā╗»┐╔─▄╩Ūę╗ĘNRNA╔·╬’īW(xu©”)ą▐’ŚĄ─äė(d©░ng)æB(t©żi)ūā╗»ģóöĄ(sh©┤)ĪŻą▐’Ś┐╔ęįė░ĒæmRNAĄ─ĮY(ji©”)śŗ(g©░u)Ż¼║═Ż©╗“Ż®ī”(du©¼)RBPsĄ─šą─╝ĪŻ╚╬║╬╠ž╩Ō╬╗³c(di©Żn)Ą─ą▐’ŚŻ¼Ģ■(hu©¼)ī¦(d©Żo)ų┬═¼ę╗mRNA╚║¾wāHāHė╔ė┌ĮY(ji©”)śŗ(g©░u)╗“╩ŪĮY(ji©”)║ŽreaderĄ─▓╗═¼Ż¼Ęų×ķā╔éĆ(g©©)mRNAüå╚║ĪŻę“┤╦Ż¼Ė─ūāą▐’ŚĄ─╗»īW(xu©”)ėŗ(j©¼)┴┐öĄ(sh©┤)Ż¼┐╔─▄╩Ū═¼ę╗éĆ(g©©)RNA▐D(zhu©Żn)õø▒Šąą╩╣▓╗═¼╣”─▄Ą─ę╗ĘNÖC(j©®)ųŲĪŻ─┐Ū░╝▒ąĶ┐╔ęįÖz£yą▐’Ś╗»īW(xu©”)ėŗ(j©¼)┴┐öĄ(sh©┤)Ą─Ė▀═©┴┐ĘĮĘ©Ż¼ęįĻU├„▒Ēė^▐D(zhu©Żn)õøĮMīW(xu©”)į┌▀@ę╗ĘĮ├µĄ─å¢Ņ}[8]ĪŻ
Arraystar mRNA&lncRNA▒Ēė^ė^▐D(zhu©Żn)õøĮMąŠŲ¼ĮY(ji©”)║Ž┴╦ļp¤╔╣Ō═©Ą└ąŠŲ¼╝╝ąg(sh©┤)┼cRNAą▐’Ś├Ōę▀╣▓│┴ĄĒ╝╝ąg(sh©┤)Ż¼į┌▐D(zhu©Żn)õø▒Š╦«ŲĮī”(du©¼)RNAą▐’Ś▀M(j©¼n)ąąČ©┴┐Öz£yĪŻČ©┴┐Ą─▒Ēė^▐D(zhu©Żn)õøĮMłDūV┐╔×ķRNAą▐’Śš{(di©żo)┐žčąŠ┐╠ß╣®ųžę¬ą┼ŽóĪŻ
┤╦═ŌŻ¼▀ĆėąArraystar small RNAą▐’ŚąŠŲ¼Ż¼┐Ą│╔╠ß╣®Ą─ŽÓĻP(gu©Īn)╝╝ąg(sh©┤)Ę■äš(w©┤)į┌å╬ÅłąŠŲ¼┐╔Č©┴┐miRNAŻ¼pre-miRNA║═tRNAč▄╔·Ą─small RNAŻ©tsRNAŻ¼░³└©tRFŻ”tiRNAŻ®Ą─ēA╗∙ą▐’ŚĪŻ┐╔Öz£yĄ─ą▐’Ś░³└©Ż║8-č§┤·°BÓč▀╩Ż©o8GŻ®Ż¼7-╝ū╗∙°B▄šŻ©m7GŻ®Ż¼N6-╝ū╗∙Ž┘▄šŻ©m6AŻ®Ż¼╝┘─“▄šŻ©”ĘŻ®╗“5-╝ū╗∙░¹▄šŻ©m5CŻ®ĪŻ
╦„╚Ī┐Ą│╔╔·╬’Arraystar ▒Ēė^▐D(zhu©Żn)õøĮMąŠŲ¼
ó┌ę║ŽÓ╔½ūV-┘|(zh©¼)ūVĘ©
LC-MS ╩Ūę╗ĘNÅŖ(qi©óng)┤¾Ą─╝╝ąg(sh©┤)Ż¼─▄ęį│÷╔½Ą─ņ`├¶Č╚║═Ė▀╠ž«ÉąįÖz£yą▐’Ś▀^Ą─RNA║╦▄šĪŻ╚╗Č°Ż¼į┌£y┴┐ mRNA ą▐’ŚĄ─▒│Š░Ž┬Ż¼ėąā╔éĆ(g©©)ĻP(gu©Īn)µIŽ▐ųŲĪŻ
╩ūŽ╚Ż¼LC-MS ▀M(j©¼n)ąą╬╗³c(di©Żn)╠žČ©Öz£yĄ──▄┴”ėąŽ▐ĪŻį┌ mRNA Ą─▒│Š░Ž┬Ż¼Ų∙Į±×ķų╣Ż¼MS āH▀mė├ė┌═Ļ╚½Ž¹╗»Ą─║╦▄šŻ¼ę“┤╦─▄ē“╣└ėŗ(j©¼)śėŲĘųąą▐’ŚĄ─Ī░š¹¾wĪ▒╦«ŲĮŻ¼Ą½┼┼│²┴╦īóįōą▐’ŚĘų┼õĄĮå╬éĆ(g©©)╬╗³c(di©Żn)Ą─┐╔─▄ąįĪŻ«ö(d©Īng)æ¬(y©®ng)ė├ė┌▓┐ĘųŽ¹╗»Ą─ RNA ╣č║╦▄š╦ßĢr(sh©¬)Ż¼LC-MS įŁät╔Ž▀Ć┐╔ęį╠ß╣®╬╗³c(di©Żn)╠žČ©ą┼ŽóĪŻ╚╗Č°Ż¼▀@ĘNĘų╬÷═©│ŻąĶę¬öĄ(sh©┤)╩«ĄĮöĄ(sh©┤)░┘╝{┐╦Ą─ų┴╔┘▓┐Ęų╝ā╗»Ą─ĘųūėŻ¼▀@ī”(du©¼)ė┌ mRNA üĒšf╩Ū▓╗¼F(xi©żn)īŹ(sh©¬)Ą─Ż¼ę“?y©żn)ķ╦³Š▀ėąĖ▀Č╚«É┘|(zh©¼)ąį║═Ą═žSČ╚ Ą─╠žąįĪŻ
Ųõ┤╬Ż¼┼c tRNA ║═ rRNA ŽÓ▒╚Ż¼mRNA ųąą▐’ŚĄ─ŽÓī”(du©¼)╦«ŲĮįĮĄ═Ż¼ę“┤╦ĮŌßī LC-MS ĮY(ji©”)╣¹ūāĄ├įĮüĒįĮ└¦ļyĪŻ╝┤╩╣üĒūįĖ▀Č╚▒Ē▀_(d©ó)Ą─ tRNA ║═ rRNA Ą─Ą═╦«ŲĮ╬█╚ŠŻ©ė└▀h(yu©Żn)¤oĘ©═Ļ╚½▒▄├ŌŻ¼▒M╣▄╦³éā┐╔ęįį┌ę╗Č©│╠Č╚╔Ž╗∙ė┌ tRNA ╠ž«Éąį╗“ rRNA ╠ž«Éąį║╦▄š▀M(j©¼n)ąą╣└ėŗ(j©¼)Ż®Ż¼ę▓┐╔─▄ī¦(d©Żo)ų┬ī”(du©¼)mRNAą▐’Ś╦«ŲĮĄ─Ė▀╣└ĪŻm6A ╩Ū MS ÅVĘ║▀M(j©¼n)ąąĄ─Ą┌ę╗éĆ(g©©)mRNA ą▐’ŚĘų╬÷Ż¼į┌▀@ĘĮ├µ╩®╝ėĄ─Ž▐ųŲ▌^╔┘Ż¼ę“?y©żn)ķ╦³į?SPAN lang=EN-US> mRNA ųąĖ▀Č╚žSĖ╗Ż¼Ą½į┌ tRNA ųą▓╗┤µį┌Ż¼▓óŪęāH┤µį┌ė┌ rRNA ųąĄ─ā╔éĆ(g©©)╬╗³c(di©Żn)Ż©ę“┤╦Ż¼╬█╚Š┐╔─▄ī¦(d©Żo)ų┬Ą═╣└īŹ(sh©¬)ļH m6A ╦«ŲĮŻ®ĪŻ
ŽÓ▒╚ų«Ž┬Ż¼Į^┤¾ČÓöĄ(sh©┤)ĘŪ m6A ▒Ēė^▐D(zhu©Żn)õøĮMį┌ mRNA ųąČ╝ĘŪ│Ż║▒ęŖŻ¼į┌ tRNA ║═ rRNA1 ųąĖ³×ķŲš▒ķŻ¼▀@ć└(y©ón)ųžŽ▐ųŲ┴╦ LC-MS ┴╦ĮŌŲõžSČ╚║═äė(d©░ng)æB(t©żi)Ą──▄┴”ĪŻį┌ę╗ą®ŪķørŽ┬Ż¼═©▀^ LC-MS ╣└ėŗ(j©¼)Ą─ą▐’ŚžSČ╚Ż©╗∙ė┌įōą▐’Ś▒╗šJ(r©©n)×ķį┌ mRNA ųąęįĖ▀╦«ŲĮ┤µį┌Ż®║═╗∙ė┌╗∙ę“ĮMĘĮĘ©Ż©═©│Ż¤oĘ©Öz£y mRNA ųąĄ─┤¾┴┐ą▐’ŚŻ®Ż╗ę“┤╦Ż¼▀@ą®▓Ņ«ÉĄ─Ė∙į┤┐╔─▄į┌ė┌ mRNA ▓┐ĘųųąĄ─ tRNA ╗“ rRNA ╬█╚Š╬’ĪŻ┴Ēę╗ĘN┐╔─▄ąį╩Ū─│ą®ą▐’Ś╩Ū RNA ōpé¹Ą─ĮY(ji©”)╣¹Ż¼▀@┐╔─▄ī¦(d©Żo)ų┬║╦▄š╦ߥ─═ķ╗∙╗»║═č§╗»ĪŻ▀@ĘNļSÖC(j©®)╔ó▓╝į┌š¹éĆ(g©©)▐D(zhu©Żn)õøĮMųąĄ─ĘŪ├Ė┤┘ą▐’Ś┐╔ęį═©▀^ LC-MS Öz£yĄĮŻ¼Ą½▓╗─▄═©▀^╗∙ę“ĮMĘĮĘ©Öz£yĄĮŻ¼▀@ąĶę¬į┌╠žČ©╬╗³c(di©Żn)Ęe└█ą┼╠¢(h©żo)ĪŻ
ó█╗∙ė┌£yą“Ą─ĘĮĘ©
į┌▀^╚ź╩«─ĻųąŻ¼╗∙ė┌Ė▀═©┴┐£yą“Ęų╬÷Ė„ĘNą▐’ŚęčĮø(j©®ng)│╔×ķ┴╦įōŅI(l©½ng)ė“▀M(j©¼n)▓ĮĄ─ĻP(gu©Īn)µI“ī(q©▒)äė(d©░ng)┴”ĪŻ▀@ą®ĘĮĘ©┐╔ęįĖµų¬ mRNA ųąą▐’ŚĄ─┤µį┌╝░Ųõį┌▐D(zhu©Żn)õøĮMųąĄ─Š½┤_╬╗ų├ĪŻ
┼c LC-MS ŽÓ▒╚Ż¼įŁät╔Žå╬éĆ(g©©)╣żū„┴„│╠Š═┐╔ęį½@ų¬╦∙ėąą▐’ŚĄ─žSČ╚Ż¼Č°╩╣ė├╗∙ę“ĮMĘĮĘ©ūR(sh©¬)äeą▐’ŚąĶę¬×ķ├┐éĆ(g©©)ą▐’Śķ_░l(f©Ī)īŻė├Ūę¬Ü(d©▓)╠žĄ─╣żū„┴„│╠ĪŻ▀@ĘNĘĮĘ©ąĶę¬┐╦Ę■Ą─Ė∙▒Š╠¶æ(zh©żn)╩ŪŻ¼┤¾ČÓöĄ(sh©┤)ą▐’Śį┌ś╦(bi©Īo)£╩(zh©│n)£yą“ųą╩ŪĪ░▓╗┐╔ęŖĄ─Ī▒Ż¼ę▓Š═╩ŪšfŻ¼╦³éā?c©©)┌ī?SPAN lang=EN-US> RNA ─µ▐D(zhu©Żn)õø×ķ cDNA ║¾▓╗Ģ■(hu©¼)┴¶Ž┬╚╬║╬║██EŻ¼▀@╩Ū╗∙ė┌ Illumina Ą─ś╦(bi©Īo)£╩(zh©│n)£yą“Ą─Ž╚øQ▓Į¾EĘĮĘ©ĪŻę“┤╦Ż¼┐ŲīW(xu©”)╝ęéāęčĮø(j©®ng)ķ_░l(f©Ī)┴╦ę╗╠ūĘĮĘ©üĒ│╩¼F(xi©żn)▓╗═¼Ą─┐╔ęŖą▐Ė─ĪŻī”(du©¼)ė┌ m6AŻ¼ų„ę¬Öz£yĘĮĘ©ę└┘ćė┌┐╣ m6A ┐╣¾wŻ¼▀@ą®┐╣¾wė├ė┌▀xō±ąį├Ōę▀│┴ĄĒ║¼ėą m6A Ą─Č╠ RNA Ų¼Č╬ĪŻ
▒M╣▄├Ōę▀│┴ĄĒ╩Ūę╗ĘN═©ė├ĘĮĘ©Ż¼įŁät╔Ž┐╔ęįė├ė┌Öz£y╚╬║╬ą▐’ŚŻ¼Ą½īŹ(sh©¬)ļH╔ŽŻ¼╗∙ė┌┐╣¾wĄ─ĘĮĘ©į┌└LųŲĘŪ m6A ▒Ēė^▐D(zhu©Żn)õøĮMĄ─łDūVĘĮ├µĄ─ą¦ė├ŽÓ«ö(d©Īng)ėąŽ▐ĪŻį°Įø(j©®ng)蹊┐╚╦åTćLįć╩╣ė├╦³üĒ└LųŲ m1A ╗“ ac4C36 Ą─łDūVŻ¼¼F(xi©żn)į┌▒╗šJ(r©©n)×ķĢ■(hu©¼)ī¦(d©Żo)ų┬┤¾┴┐╝┘Ļ¢ąį╬╗³c(di©Żn)Ż¼▀@┐╔─▄╩Ūė╔ė┌┐╣¾wĮ╗▓µĘ┤æ¬(y©®ng)įņ│╔Ą─ĪŻ┐╣ m6A ┐╣¾wę▓┤µį┌▀@ĘNĮ╗▓µĘ┤æ¬(y©®ng)ąįŻ╗╚╗Č°Ż¼Ķbė┌ m6A Ą─žSČ╚ŽÓī”(du©¼)▌^Ė▀Ż¼ą┼įļ▒╚╚į╚╗┐╔┐žĪŻŽÓ▒╚ų«Ž┬Ż¼ęį▌^Ą═öĄ(sh©┤)┴┐╝ē(j©¬)Ą─žSČ╚┤µį┌Ą─ą▐’Śī¦(d©Żo)ų┬ĘŪ╠ž«Éąį┼c╠ž«ÉąįĮY(ji©”)║Ž╩┬╝■Ą─▒╚┬╩╝▒äĪį÷╝ėĪŻ
ę“┤╦Ż¼ī”(du©¼)ė┌Į^┤¾ČÓöĄ(sh©┤)ĘŪ m6A ▒Ēė^▐D(zhu©Żn)õøĮMŻ¼ęčĮø(j©®ng)ķ_░l(f©Ī)│÷└¹ė├ą▐’ŚēA╗∙Ą─¬Ü(d©▓)╠ž╗»īW(xu©”)ąį┘|(zh©¼)╩╣╦³éā?c©©)┌─µ▐D(zhu©Żn)õø║¾┐╔ęŖĄ─ĘĮĘ©ĪŻ▀@╩Ū═©▀^▓╗═¼Ą─▓▀┬įīŹ(sh©¬)¼F(xi©żn)Ą─Ż¼└²╚ńŻ¼═©▀^Ė─ūāą▐’ŚēA╗∙╔ĒĘ▌Ą─ą▐’Ś╠ž«Éąį╗»īW(xu©”)╬’┘|(zh©¼)╗“═©▀^īó┤¾┴┐Üł╗∙╠žČ©▀BĮėĄĮą▐’ŚēA╗∙╔Žī¦(d©Żo)ų┬─µ▐D(zhu©Żn)õø▀^│╠ųą▀^įńĮž?c©ói)ÓŻ©ęŖŽ┬▒ĒŻ®ĪŻ▀@ą®īŹ(sh©¬)“×(y©żn)ĘĮ░Ė═©│ŻĢ■(hu©¼)ī¦(d©Żo)ų┬ą▐Ė─╬╗³c(di©Żn)Ą─Õe(cu©░)┼õ╗“╚▒╩¦Ż¼╗“š▀ī¦(d©Żo)ų┬ūx╚ĪĄ─Ī░ČčĘeĪ▒Ż¼▀@ą®ūx╚Ī▀xō±ąįĄžį┌╠žČ©╬╗ų├ķ_╩╝╗“ĮY(ji©”)╩°Ż¼╚╗║¾┐╔ęįÅ─ųą═ŲöÓ│÷ą▐Ė─Ą─┤µį┌ĪŻ┤╦ŅÉ╗»īW(xu©”)╗∙ę“ĮMīW(xu©”)ĘĮĘ©Ą─ų„ę¬ā×(y©Łu)ä▌į┌ė┌Ųõ│÷╔½Ą─Īó═©│Ż×ķå╬║╦▄š╦ߥ─Ęų▒µ┬╩Ż¼ęį╝░╦³éā─▄ē“╠ß╣®ą▐’Ś╦«ŲĮĄ─ŽÓī”(du©¼)Č©┴┐Ż¼ėąĢr(sh©¬)╔§ų┴╩ŪĮ^ī”(du©¼)Č©┴┐[1]ĪŻ
▒╚╚ń┐Ą│╔╔·╬’Į©┴ó┴╦ę╗ĘNĖ▀ņ`├¶Ą─Į^ī”(du©¼)Č©┴┐Ą─īŹ(sh©¬)Ģr(sh©¬)¤╔╣ŌČ©┴┐PCRČ©┴┐Ęų╬÷ĘĮĘ©Ż¼┐╔ęįį┌å╬║╦▄š╦ßĘų▒µ┬╩╦«ŲĮ╔Ž£╩(zh©│n)┤_Öz£yRNAųąm6Aą▐’Ś╬╗³c(di©Żn), ▓óī”(du©¼)╬┤ų¬─Ż░Õ▀M(j©¼n)ąąČ©┴┐Ęų╬÷ĪŻįōĘĮĘ©┐╔│╔╣”Ąžæ¬(y©®ng)ė├ė┌īŹ(sh©¬)ļH╔·╬’śėŲĘųąm6Aą▐’ŚĄ─Š½┤_Öz£yŻ¼╝┤╩╣╩ŪĄ═žSČ╚RNAĄ─śė▒ŠĪŻ
╦„╚Ī┐Ą│╔Arraystar m6Aå╬ēA╗∙Ęų▒µ┬╩ąŠŲ¼╝╝ąg(sh©┤)┘Y┴Ž>>
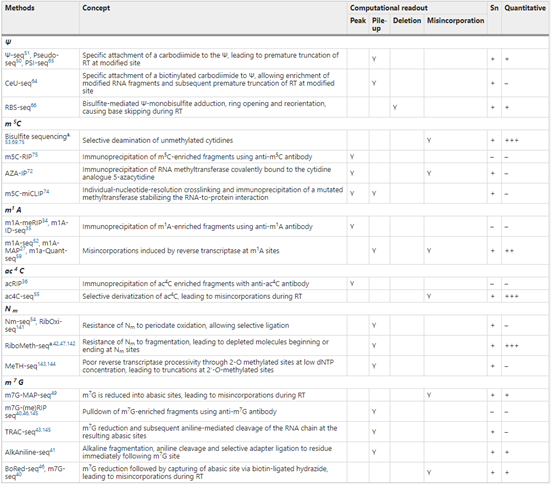
Į³─ĻüĒę▓ėąę╗ą®ą┬Ą─£yą“╝╝ąg(sh©┤)Ļæ└m(x©┤)│÷¼F(xi©żn)Ż¼▒╚╚ń2017 ─ĻŻ¼▒▒Š®┤¾īW(xu©”)ę┴│╔Ų„šnŅ}ĮM║═ęį╔½┴ą╬║┤─┬³čąŠ┐╦∙Ą─蹊┐łF(tu©ón)ĻĀ(du©¼)Ęųäe¬Ü(d©▓)┴ół¾(b©żo)Ą└┴╦å╬ēA╗∙Ęų▒µ┬╩╦«ŲĮÖz£y m1A Ą─ą┬ĘĮĘ©Ī¬Ī¬m1A-MAP ║═m1A-SeqĪŻ2019 ─ĻŻ¼ųź╝ėĖń┤¾īW(xu©”) Bryan CŻ« Dickinson łF(tu©ón)ĻĀ(du©¼)┼c║╬┤©łF(tu©ón)ĻĀ(du©¼)║Žū„Ż¼čąŠ┐ķ_░l(f©Ī)┴╦┐╔┐ņ╦┘▀xō±─µ▐D(zhu©Żn)õø├ĖĄ─▀M(j©¼n)╗»ŲĮ┼_(t©ói)Ż¼▓ó└¹ė├įōŲĮ┼_(t©ói)蹊┐ mRNA ╔ŽĄ─ m1A ą▐’ŚŻ¼īŹ(sh©¬)¼F(xi©żn)┴╦å╬ēA╗∙╦«ŲĮ m1A Ą─Öz£yĪŻ
┴Ē═ŌŻ¼ų▒ĮėĄ─RNA╝{├ū┐ū£yą“ę▓éõ╩▄ĻP(gu©Īn)ūóŻ¼RNA▒Ēė^▀zé„īW(xu©”)Ą─Ž╚“ī(q©▒)╚╦╬’Īó┐Ą─╬Ā¢┤¾īW(xu©”)Ą─Christopher Mason▒Ē╩ŠŻ¼Ī░▀^╚źŻ¼╬ęéā═©│Ż╩Ū└¹ė├┐╣¾w╗“╗»īW(xu©”)Ęų╬÷üĒ═ŲöÓRNAĄ─ą▐’ŚĀŅæB(t©żi)ĪŻų▒ĄĮūŅĮ³Ż¼╬ęéā▓┼ķ_╩╝▀M(j©¼n)ąąų▒ĮėĄ─RNA£yą“ĪŻ└¹ė├╝{├ū┐ū£yą“āxŻ¼╬ęéā─▄ē“ų▒Įė£yą“RNAŻ¼Č°▓╗ąĶę¬─µ▐D(zhu©Żn)õøĪŻ╬ęéāĄ┌ę╗┤╬ų▒Įė£yČ©RNAą▐’ŚŻ║š¹éĆ(g©©)Ęųūė┤µį┌──ą®ą▐’ŚŻ¼ėų┤µį┌──ą®«Éśŗ(g©░u)¾wĪŻĪ▒
╦∙ęį┐╔ęįšfŻ¼╝{├ū┐ū£yą“─▄═Ļš¹£yą“Ż¼ūx│÷RNAą“┴ąĄ─m6A,m5Cą▐’ŚŻ©ę╗░Ń╩Ūūx╚Īå╬Ęųūėą“┴ąĢr(sh©¬)▒╚¤oą▐’ŚēA╗∙ėąčė▀tŻ®Ż¼ūxķLā×(y©Łu)ä▌╩╣ų«ī”(du©¼)┐╔ūā╝¶ŪąčąŠ┐║═«Éśŗ(g©░u)¾wĖ³ėąā×(y©Łu)ä▌Ż¼▓╗ė├Ų┤ĮėŻ¼ę▓▓╗╚▌ęūüG╩¦║▒ęŖĘųūėŻ¼Ą½╩ŪŠ═╩Ū┘M(f©©i)ė├▒╚▌^┘FĪŻ
į┌ūŅĮ³Ą─ę╗Ų¬¤ß³c(di©Żn)╬─š┬Ī░The Architecture of SARS-CoV-2 Transcriptome SARS-CoV-2Ī▒ųąŻ¼čąŠ┐╚╦åTŠ═ė├MinION╝{├ū┐ū£yą“āx▀M(j©¼n)ąą┴╦DRS£yą“Ż¼½@Ą├┴╦879,679éĆ(g©©)readsŻ©1.9 GbŻ®Ż¼ĮY(ji©”)╣¹▓╗āHūC├„┴╦▓ĪČŠ▐D(zhu©Żn)õø▒Šš╝ō■(j©┤)ų„ī¦(d©Żo)Ąž╬╗Ż¼Č°Ūę╝{├ū┐ūDRS╗∙ė┌RNAĄ─å╬ĘųūėÖz£yŻ¼╠ß╣®┴╦¬Ü(d©▓)╠žĄ─ÖC(j©®)Ģ■(hu©¼)üĒÖz▓ķå╬éĆ(g©©)RNAĘųūėĄ─ČÓéĆ(g©©)▐D(zhu©Żn)õøĮM╠žš„Ż¼ĮŌ╬÷┴╦ą┬╣┌▓ĪČŠRNAą▐’Ś╠ž³c(di©Żn)[9]ĪŻ
óŲ─┐Ū░ęčų¬Ą─RNA▒Ēė^▐D(zhu©Żn)õøą▐’ŚŻ║
m6A
m6A╩Ūšµ║╦╔·╬’mRNA╔Ž║¼┴┐ūŅžSĖ╗Ą─╗»īW(xu©”)ą▐’ŚŻ¼ė╔╝ū╗∙▐D(zhu©Żn)ęŲ├ĖÅ═(f©┤)║Ž╬’Ż©░³║¼METTL3Ż¼METTL14Ż¼WTAPŻ¼KIAA1429Ż¼RBM15Ż¼RBM15BŻ®┤▀╗»«a(ch©Żn)╔·Ż¼┐╔▒╗╚ź╝ū╗∙╗»├ĖALKBH5╗“FTO╚ź│²ĪŻ─┐Ū░ęč░l(f©Ī)¼F(xi©żn)┴╦ČÓĘN╠ž«ÉąįūR(sh©¬)äem6A╬╗³c(di©Żn)Ą─Ą░░ū╗“Å═(f©┤)║Ž╬’Ż¼░³└©YTH╝ęūÕĄ░░ūŻ©YTHDF1-3Ż¼YTHDC1Ż®Īó▐D(zhu©Żn)õøŲ╩╝Å═(f©┤)║Ž╬’eIF3Īó║╦╠Ū║╦Ą░░ūŻ©HNRNPA2B1Ż¼HNRNPCŻ®ęį╝░RNAĮY(ji©”)║ŽĄ░░ūSRSF2ĪŻm6Aų„ę¬Ęų▓╝į┌ĮKų╣├▄┤aūėĖĮĮ³║═3Ī»UTRģ^(q©▒)Ż¼ė░ĒæRNA┼õī”(du©¼)ĪóĖ─ūāRNAČ■╝ē(j©¬)ĮY(ji©”)śŗ(g©░u)Īó╗“▒╗Ą░░ūų▒ĮėūR(sh©¬)äeŻ¼▀M(j©¼n)Č°š{(di©żo)┐žmRNAĄ─│╔╩ņĪó┐╔ūā╝¶ĮėĪóĘĆ(w©¦n)Č©ąį║═ĘŁūg▀^│╠ĪŻ┼cA-U┼õī”(du©¼)ŽÓ▒╚Ż¼m6A-U┼õī”(du©¼)▌^▓╗ĘĆ(w©¦n)Č©Ż¼ę²░l(f©Ī)RNAā╚(n©©i)▓┐ļpµ£ĮŌą²┼cČ■╝ē(j©¬)ĮY(ji©”)śŗ(g©░u)▐D(zhu©Żn)ūāĪŻm6A│Żį┌ļpµ£┼cå╬µ£Ą─▀^Č╔ģ^(q©▒)ė“Čč»BŻ¼į÷ÅŖ(qi©óng)RNA▐D(zhu©Żn)ūā║¾śŗ(g©░u)Ž¾Ą─ĘĆ(w©¦n)Č©ąįĪŻ╚ź╝ū╗∙╗»ät┐╔ęį╩╣mRNA╗ųÅ═(f©┤)įŁüĒśŗ(g©░u)Ž¾ĪŻ▀@ĘNśŗ(g©░u)Ž¾▐D(zhu©Żn)ūā┐╔─▄ī¦(d©Żo)ų┬mRNA┼c▓╗═¼Ą░░ūĄ─ŽÓ╗źū„ė├Ė─ūāŻ¼Å─Č°«a(ch©Żn)╔·▓╗═¼Ą─╔·╬’īW(xu©”)ą¦æ¬(y©®ng)ĪŻm6A─▄ų▒Įė▒╗╠žČ©Ą░░ūĄ─╩Ķ╦«ĮY(ji©”)śŗ(g©░u)ė“╦∙ūR(sh©¬)äeĪŻ▒╚╚ńŻ¼YTH╝ęūÕĄ░░ū┐╔ęį╠ž«ÉąįĄ─ūR(sh©¬)äem6AŻ¼╠žäe╩ŪGGm6ACU▒Ż╩žą“┴ąĪŻŲõ│╔åTų«ę╗YTHDC1ūR(sh©¬)äe▓óĮY(ji©”)║Žm6AŻ¼š{(di©żo)┐ž░ąŽ“mRNAĄ─┐╔ūā╝¶ĮėĪŻČ°┴Ēę╗│╔åTYRHDF2┼cm6AĮY(ji©”)║Ž║¾Ż¼šą─╝CCR4-NOTÅ═(f©┤)║Ž╬’Ż¼┤┘▀M(j©¼n)░ąŽ“RNAĄ─ĮĄĮŌĪŻį┌UV▌Ś╔õ╗“¤ßą▌┐╦Ę┤æ¬(y©®ng)ųąŻ¼▐D(zhu©Żn)õøŲ╩╝Å═(f©┤)║Ž╬’eIF3┼c5Ī»UTRģ^(q©▒)Ą─m6AĮY(ji©”)║ŽŻ¼┤┘▀M(j©¼n)├▒ĘŪę└┘ćĄ─ĘŁūg▀^│╠ĪŻŠÄ┤aģ^(q©▒)Ą─m6A┐╔▒╗SRSF2ūR(sh©¬)äeŻ¼ģó┼cų¼ĘŠ╔·│╔Ą─š{(di©żo)┐žĪŻį┌╣¹ŽēųąŻ¼YTHDC1═¼į┤Ą░░ūūR(sh©¬)äeąįäeų┬╦└mRNA╔ŽĄ─m6A╬╗³c(di©Żn)Ż¼š{(di©żo)┐žŲõ┐╔ūā╝¶ĮėŻ¼Å─Č°┐žųŲ╣¹ŽēĄ─ąįäeĪŻ╚ńŪ░╦∙╩÷Ż¼m6AĮķī¦(d©Żo)Ą─mRNAĘĆ(w©¦n)Č©ąįš{(di©żo)╣Ø(ji©”)ę▓ī”(du©¼)Ė╔╝Ü(x©¼)░¹Ęų╗»┼c╔·╬’╣Ø(ji©”)┬╔Ģr(sh©¬)ńŖ┐žųŲ╩«Ęųųžę¬ĪŻ┤╦═ŌŻ¼m6Aę▓─▄═©▀^ė░ĒæmRNA┼ctRNAĘ┤├▄┤aūėĄ─┼õī”(du©¼)╦┘┬╩┼c▒ŻšµČ╚Ż¼Å─Č°ė░ĒæĘŁūgčė╔ņĪŻ
蹊┐ĘĮĘ©ĘĮ├µŻ¼Arraystar m6Aå╬ēA╗∙Ęų▒µ┬╩ąŠŲ¼└¹ė├RNA├ĖMazFī”(du©¼)m6A├¶ĖąĄ─╠žąįŻ¼┐╔į┌å╬ēA╗∙Ęų▒µ┬╩╦«ŲĮČ©╬╗m6A╬╗³c(di©Żn)Ż¼▓óī”(du©¼)Ųõ▀M(j©¼n)ąąČ©┴┐ĪŻįōąŠŲ¼Š▀ėąŲõ╦³¼F(xi©żn)┤µ╝╝ąg(sh©┤)ļyęįŲ¾╝░Ą─ā×(y©Łu)ä▌Ż¼╩Ū蹊┐m6Aäė(d©░ng)æB(t©żi)ūā╗»Īó╔·╬’īW(xu©”)╣”─▄ęį╝░╝▓▓ĪŽÓĻP(gu©Īn)ąįĄ─ÅŖ(qi©óng)┤¾╣żŠ▀ĪŻ
m1A
m1A╩Ūą┬Į³░l(f©Ī)¼F(xi©żn)Ą─┐╔─µĄ─▒Ēė^▐D(zhu©Żn)õøą▐’ŚŻ¼─▄▒╗RNAą▐Å═(f©┤)├ĖALKBH3╚ź│²Ż¼─┐Ū░╔ą╬┤░l(f©Ī)¼F(xi©żn)├„┤_Ą─m1Aą▐’Ś├Ė┼cą▐’ŚūR(sh©¬)äeĄ░░ūĪŻ┼cm6A▓╗═¼Ż¼m1AĄ─▒Ē▀_(d©ó)žSČ╚▌^Ą═Ż¼ų„ę¬Ęų▓╝į┌mRNAĄ─5Ī»UTRģ^(q©▒)Ż¼┐╔─▄ģó┼cš{(di©żo)╣Ø(ji©”)ĘŁūgŲ╩╝▀^│╠ĪŻm1A─▄═Ļ╚½ūĶų╣Watson-Crick┼õī”(du©¼)Ż¼ę²ŲRNAļpµ£ĮŌą²Ż¼▓ó┤┘▀M(j©¼n)RNA-Ą░░ūĄ─ņoļŖŽÓ╗źū„ė├╗“RNA┐╔ūāČ■╝ē(j©¬)ĮY(ji©”)śŗ(g©░u)Ą─ą╬│╔ĪŻm1AĄ─╔·╬’īW(xu©”)╣”─▄╚įī┘╬┤ų¬ĪŻėąčąŠ┐░l(f©Ī)¼F(xi©żn)Ż¼į┌¤ßą▌┐╦╗“ĀIB(y©Żng)ģTĘ”Ą╚ē║┴”Śl╝■Ž┬Ż¼╝Ü(x©¼)░¹ā╚(n©©i)Ą─m1A▒Ē▀_(d©ó)╦«ŲĮ╔Ž╔²Ż¼┐╔─▄╩Ū═©▀^┤┘▀M(j©¼n)├▒ę└┘ćąįĘŁūgŻ¼ģó┼c╝Ü(x©¼)░¹Ą─æ¬(y©®ng)╝żĘ┤æ¬(y©®ng)ĪŻ
m5C┼chm5C
m5CÅVĘ║Ęų▓╝ė┌tRNA┼crRNA╔ŽŻ¼Š▀ėąĘĆ(w©¦n)Č©tRNAČ■╝ē(j©¬)ĮY(ji©”)śŗ(g©░u)Īóė░ĒæĘ┤├▄┤aūėŁh(hu©ón)śŗ(g©░u)Ž¾ĪóŠS│ųrRNAĘŁūg▒ŻšµĄ╚╣”─▄ĪŻą┬Į³Ą─RNA£yą“ĮY(ji©”)╣¹░l(f©Ī)¼F(xi©żn)Ż¼mRNAĄ─ŠÄ┤aģ^(q©▒)┼cĘŪŠÄ┤aģ^(q©▒)╔Ž┤µį┌8000ČÓéĆ(g©©)m5C╬╗³c(di©Żn)Ż¼Č°ŪęŽÓ«ö(d©Īng)ę╗▓┐Ęų╬╗³c(di©Żn)╝»ųąį┌5Ī»UTR┼c3Ī»UTRģ^(q©▒)ĪŻm5C┐╔ė╔╝ū╗∙▐D(zhu©Żn)ęŲ├ĖNSUN2╗“TRDMT1┤▀╗»ą╬│╔Ż¼▒╗ļp╝ėč§├ĖTETč§╗»ą╬│╔hm5CĪŻhm5C┐╔─▄Įø(j©®ng)▀^▀M(j©¼n)ę╗▓Įč§╗»ą╬│╔f5CŻ¼▀M(j©¼n)Č°ūā╗ž░¹ÓūÓż║╦▄šŻ©CŻ®ĪŻm5C▓╗ė░ĒæēA╗∙┼õī”(du©¼)Ż¼Ą½┐╔─▄į÷ÅŖ(qi©óng)ēA╗∙Čč»Bęį╝░RNA┼cĄ░░ūĄ─╩Ķ╦«ū„ė├ĪŻm5CŠ▀ėąČÓĘN╔·╬’īW(xu©”)╣”─▄ĪŻp16 mRNAį┌▒╗NSUN2├Ė╠Ē╝ėm5Cą▐’Ś║¾Ż¼ŲõĮĄĮŌ▒╗ęųųŲŻ¼ĘĆ(w©¦n)Č©ąįį÷ÅŖ(qi©óng)ĪŻį┌╝Ü(x©¼)░¹ų▄Ų┌ųąŻ¼NSUN2Ą─▒Ē▀_(d©ó)╩▄ĄĮŠ½├▄š{(di©żo)┐žĪŻNSUN2┐╔į┌CDK1 3Ī»UTRģ^(q©▒)╠Ē╝ėm5Cą▐’ŚŻ¼┤┘▀M(j©¼n)ŲõĘŁūgŻ╗═¼Ģr(sh©¬)į┌CDKN1BĄ─5Ī»UTRģ^(q©▒)╠Ē╝ėm5Cą▐’ŚŻ¼ęųųŲCDKN1BĄ─ĘŁūgĪŻČ■š▀╣▓═¼ū„ė├Ż¼į÷ÅŖ(qi©óng)╝Ü(x©¼)░¹Ą─į÷ų│─▄┴”ĪŻm5C▀Ć┼c╦ź└ŽŽÓĻP(gu©Īn)╗∙ę“Ą─ĘŁūg┐žųŲėąĻP(gu©Īn)Ż¼▀^▒Ē▀_(d©ó)NSUN2┐╔ęįčėŠÅÅ═(f©┤)ųŲąį╦ź└ŽĄ─░l(f©Ī)╔·ĪŻū„×ķm5CĄ─č§╗»«a(ch©Żn)╬’Ż¼hm5Cę▓─▄į÷ÅŖ(qi©óng)ĘŁūgą¦┬╩ĪŻhm5Cį┌╣¹ŽēĄ──Xųą▒Ē▀_(d©ó)┴┐║▄Ė▀Ż¼┐╔─▄ģó┼c╣¹ŽēĄ──X▓┐░l(f©Ī)ė²ĪŻ
Pseudouridine (”ū)
╝┘─“ÓūÓż║╦▄š”ūŻ¼│Ż▒╗│╔×ķĄ┌╬ÕŅÉ║╦▄š╦߯¼ė╔─“ÓūÓż║╦▄šŻ©UŻ®«Éśŗ(g©░u)╗»ą╬│╔ĪŻį┌╚╦Ą─╝Ü(x©¼)░¹║═ąĪ╩¾ĮM┐ŚĄ─mRNAųąŻ¼”ū/UĄ─▒╚┬╩╝s×ķ0.2-0.6%ĪŻ─“ÓūÓż┼c╝┘─“ÓūÓżĄ─«Éśŗ(g©░u)╗»Ę┤æ¬(y©®ng)ė╔PUS├Ėå╬¬Ü(d©▓)Ż¼╗“┼cH/ACA║╦╠Ū║╦Ą░░ūę╗ŲŻ¼┤▀╗»═Ļ│╔ĪŻ╝┘─“ÓūÓż─▄£p╔┘RNAśŗ(g©░u)Ž¾Ą─┐╔ūāąįŻ¼į÷ÅŖ(qi©óng)ēA╗∙┼õī”(du©¼)ĘĆ(w©¦n)Č©ąįęį╝░┼cĄ░░ūų«ķgĄ─Ą─śOąįŽÓ╗źū„ė├ĪŻ╝┘─“ÓūÓż┐╔─▄š{(di©żo)┐žmRNAĘĆ(w©¦n)Č©ąį║═╗∙ę“▒Ē▀_(d©ó)Ż¼ģó┼cĮ═─ĖĄ─¤ßą▌┐╦Ę┤æ¬(y©®ng)Ż¼Ą½Š▀¾wÖC(j©®)ųŲ─┐Ū░╔ą▓╗├„┤_ĪŻ
Inosine (I)
╝Ī▄šą▐’ŚŻ¼│ŻĘQ×ķA-to-IŠÄ▌ŗŻ¼╩ŪĖ▀Ą╚šµ║╦╔·╬’└’ūŅ│ŻęŖĄ─ę╗ĘNRNAŠÄ▌ŗĘĮ╩ĮŻ¼ė╔Ž┘▄š╦ß├ō░▒├ĖADAR═Ļ│╔ĪŻA-to-IŠÄ▌ŗų„ę¬░l(f©Ī)╔·į┌ĘŪŠÄ┤aģ^(q©▒)╗“ā╚(n©©i)║¼ūėģ^(q©▒)Ą─Aluį¬╦žā╚(n©©i)ĪŻA-to-IŠÄ▌ŗ═Ļ╚½Ė─ūā┴╦ēA╗∙┼õī”(du©¼)╠žąįŻ¼AU┼õī”(du©¼)▐D(zhu©Żn)ūā?y©Łu)?SPAN lang=EN-US>IC┼õī”(du©¼)Ż¼Å─Č°Ė─ūā╦∙ŠÄ┤aĄ─░▒╗∙╦ßĪŻ▒╚╚ńŻ¼A-to-IŠÄ▌ŗīó┤¾─X╣╚░▒╦ß╩▄¾wųąĄ─╣╚░▒§Ż░ĘųžŠÄ┤a×ķŠ½░▒╦߯¼ī¦(d©Żo)ų┬┴╦Ō}ļxūė═©═ĖąįĄ─Ė─ūāĪŻ┤╦═ŌŻ¼A-to-IŠÄ▌ŗ▀ĆŠ▀ėąĖ─ūā┐╔ūā╝¶ĮėĪóš{(di©żo)╣Ø(ji©”)miRNAĄ─«a(ch©Żn)╔·┼c╣”─▄Īóęį╝░▒O(ji©Īn)┐žŽ╚╠ņąį├Ōę▀Ę┤æ¬(y©®ng)Ą╚ū„ė├ĪŻ
ac4C
2018─ĻŻ¼├└ć°NIHĄ─Shalini OberdoerfferĮ╠╩┌į┌Cell░l(f©Ī)▒Ē蹊┐šō╬─Ż║Acetylation of Cytidine in mRNA Promotes Translation EfficiencyŻ¼╩ū┤╬Įę╩ŠmRNA╔Ž┤µį┌┤¾┴┐ac4Cą▐’ŚŻ¼▓óŪęac4Cė░ĒæmRNAĄ─ĘĆ(w©¦n)Č©ąį┼cĘŁūgą¦┬╩ĪŻ
N4-acetylcytidine (ac4C)Ż¼N4╬╗ęꧯ░¹ÓūÓżŻ¼╩Ūšµ║╦įŁ║╦╔·╬’ųą▒Ż╩žĄ─╗»īW(xu©”)ą▐’ŚŻ¼įńŲ┌蹊┐šJ(r©©n)×ķac4Cų„ę¬┤µį┌tRNA║═18S rRNA╔ŽĪŻČ°Į³Ų┌蹊┐’@╩ŠŻ¼mRNA╔Žę▓┤µį┌┤¾┴┐Ą─ac4CŻ¼ŲõžSČ╚╔§ų┴▓╗Ą═ė┌mRNAöyĦĄ─m7G├▒ūėą▐’ŚĪŻNAT10╩Ū─┐Ū░ĶbČ©Ą─╬©ę╗═¼Ģr(sh©¬)Š▀ėąęꧯ╗»├ĖĮY(ji©”)śŗ(g©░u)ė“║═RNAĮY(ji©”)║ŽĮY(ji©”)śŗ(g©░u)ė“Ą─Ą░░ūŻ¼ę“┤╦▒╗šJ(r©©n)×ķ╩ŪRNA ac4Cą▐’Ś├ĖĪŻ
ė╔┤╦Ż¼ęꧯ╗»ū„×ķę╗ŅÉą┬Ą─ mRNA ą▐’ŚŻ¼īó▒Ēė^▐D(zhu©Żn)õøĮMöU(ku©░)š╣ĄĮęčų¬ŅÉäeĄ─╝ū╗∙╗»║═«Éśŗ(g©░u)╗»ų«═ŌĪŻ▀@ą®╣żū„▀ĆöU(ku©░)š╣┴╦ mRNA ą▐’ŚĄ─ęčų¬ÄņŻ¼▓ó┤_Č©┴╦ČÓĘN╗»īW(xu©”)ą▐’ŚĄ─┤µį┌Ż¼mRNA ą▐’ŚĄ─ą┬─Ż╩Įį┌╬┤üĒ║▄┐╔─▄Ģ■(hu©¼)└^└m(x©┤)│÷¼F(xi©żn)ĪŻ▀M(j©¼n)ę╗▓Į┴╦ĮŌ▀@ą®ą▐’ŚĄ─╣”─▄Īóš{(di©żo)╣Ø(ji©”)╦³éā?c©©)┌╠žČ©▐D(zhu©Żn)õø▒Š╔ŽĄ─┤µį┌Ą─═ŠÅĮ║═ÖC(j©®)ųŲŻ¼ęį╝░╦³éā?c©©)┌Ė„ĘN╔·└Ē║═╝▓▓Ī▒│Š░Ž┬Ą─╣”─▄Ż¼īóėąų·ė┌Ė³╚½├µĄž┴╦ĮŌ▒Ēė^▐D(zhu©Żn)õøĮMį┌╗∙ę“▒Ē▀_(d©ó)ųąĄ─ū„ė├[6]ĪŻ
TGMą▐’Ś
5Ī»Č╦│¼╝ū╗∙╗»Ą─TMGą▐’Ś╩ŪūŅįń▒╗░l(f©Ī)¼F(xi©żn)Ą─ą▐’Śų«ę╗ĪŻį┌įSČÓ▒╗RNAŠ█║Ž├ĖII▐D(zhu©Żn)õøĄ─RNAųąŻ¼ė╚Ųõ╩ŪĘŪŠÄ┤aRNAŻ¼└²╚ń╝¶Įė¾wĄ─ųžę¬ĮM│╔│╔ĘųsnRNAŻ¼5Ī»Č╦Ą─m7G╝ū╗∙├▒ūėĢ■(hu©¼)▒╗│¼╝ū╗∙╗»Ż¼ą╬│╔Š▀ėą╚²éĆ(g©©)╝ū╗∙Ą─N2, N2, 7-╚²╝ū╗∙°B▄šŻ©TMGŻ®├▒ūėŻ¼ÅVĘ║┤µį┌ė┌šµ║╦╔·╬’ųąĪŻ╚²╝ū╗∙°B▄š║Ž│╔├ĖŻ©Tgs1Ż®į┌įSČÓ╔·╬’¾wųąęčĮø(j©®ng)▒╗ūCīŹ(sh©¬)╩Ū║Ž│╔TMG├▒ūėĄ─╬©ę╗╝ū╗∙├ĖŻ¼į┌▀M(j©¼n)╗»╔ŽÅ─Į═─ĖĄĮ╚╦ŅÉČ╝║▄▒Ż╩žĪŻė╔ė┌¾w═ŌīŹ(sh©¬)“×(y©żn)Ą─ŠųŽ▐ąįŻ¼TMGą▐’Śī”(du©¼)╝¶Įė¾wRNAĄ─š{(di©żo)┐žÖC(j©®)ųŲ▀Ć┤µį┌║▄ČÓ▓╗ŪÕ│■Ą─ĄžĘĮŻ¼▓óŪęį┌┤¾ČÓöĄ(sh©┤)ČÓ╝Ü(x©¼)░¹─Ż╩Į╔·╬’ųąČ╝¤oĘ©ų▒ĮėÅ─¾wā╚(n©©i)ī”(du©¼)TMGą▐’Ś▀M(j©¼n)ąąčąŠ┐Ż¼ę“┤╦Ż¼TMGą▐’ŚęčĮø(j©®ng)▒╗░l(f©Ī)¼F(xi©żn)┴╦╬Õ╩«ČÓ─ĻŻ¼Ą½╩ŪŲõį┌ČÓ╝Ü(x©¼)░¹╔·╬’ųąĄ─╣”─▄▓ó▓╗├„┤_ĪŻ
Ż©╔·╬’═©Ż║╚f╝yŻ®
ģó┐╝╬─½I(xi©żn)Ż║
1. The epitranscriptome beyond m6A,Nature Reviews Genetics volume 22, pages119©C131 (2021)
2. Epitranscriptome sequencing technologies: decoding RNA modifications,Nature Methods volume 14, pages23©C31 (2017)
3. Detecting RNA modifications in the epitranscriptome: predict and validate,Nature Reviews Genetics volume 18, pages275©C291 (2017)
4. RNA editing-dependent epitranscriptome diversity in cancer stem cells,Nature Reviews Cancer volume 17, pages381©C392 (2017)
5. RNA modifications: what have we learned and where are we headed? Nature Reviews Genetics volume 17, pages365©C372 (2016)
6. mRNA acetylation: a new addition to the epitranscriptomeŻ¼P. Cody He & Chuan He Cell Research volume 29, pages91©C92 (2019)
7. Wendy V. Gilbert, Tristan A. Bell, Cassandra Schaening. Science (2016)
8. Cole J.T. Lewis, Tao Pan, Auinash Kalsotra. Nat Rev Mol Cell Biol (2017)
9. The Architecture of SARS-CoV-2 Transcriptome,Cell,April 23Ż¼2020
╔·╬’═©╬óą┼╣½▒Ŗ╠¢(h©żo)
Į±╚šäė(d©░ng)æB(t©żi) | ╚╦▓┼╩ął÷ | ą┬╝╝ąg(sh©┤)īŻÖ┌ | ųąć°┐ŲīW(xu©”)╚╦ | įŲš╣┼_(t©ói) | BioHot | įŲųv╠├ų▒▓ź | Ģ■(hu©¼)š╣ųąą─ | ╠žār(ji©ż)īŻÖ┌ | ╝╝ąg(sh©┤)┐ņėŹ | ├Ō┘M(f©©i)įćė├
░µÖÓ(qu©ón)╦∙ėą ╔·╬’═©
Copyright© eBiotrade.com, All Rights Reserved
┬ō(li©ón)ŽĄą┼ŽõŻ║
╗øICPéõ09063491╠¢(h©żo)
