- ���(y��) ���Մ�(d��ng)�B(t��i) �˲��Ј�(ch��ng) �¼��g(sh��)��(zhu��n)�� �Ї�(gu��)�ƌW(xu��)�� ��չ�_(t��i) ���v��ֱ�� ��(hu��)չ���� �r(ji��)��(zhu��n)�� ���g(sh��)��Ӎ ���M(f��i)ԇ��

-

����ͨ��
����ץס�����Ƽ�
����(d��ng)���}��
����ͨ��
����ץס�����Ƽ�
����(d��ng)���}��
m6A�{(di��o)��(ji��)��ω�Ԅe�ֻ�����(j��ng)���� | ��������
�����w�� �� �� С �� �r(sh��)�g��2018��05��08�� ��(l��i)Դ��(li��n)������
�����]��
���� (t��ng) (t��ng)��(zhu��n)�ҽ���m6A�о��ľ��w��������m6A�{(di��o)��(ji��)��ω�Ԅe�ֻ�����(j��ng)���ܡ�
��(bi��o)�}��m6A modulates neuronal functions and sex determination in Drosophila
m6A�{(di��o)��(ji��)��ω�Ԅe�ֻ�����(j��ng)����
�l(f��)��r(sh��)�g��2016��12��
�l(f��)���ڿ���Nature
IF��40.1
�C(j��)��(g��u)����(gu��)����Ĵ�W(xu��)
1��ǰ��
RNA����ڱ��^�D(zhu��n)��{(di��o)����ʮ����Ҫ������m6A������S�ಸ���(d��ng)���ЏV�����ڡ��S����������ж�����m6A��������ˡ�С���R�~(y��)������ĸ���S���о�����������m6A���������RRACH motif��R, purine; H, non-guanine base����(n��i)������Kֹ�ܴa�Ӻ�3�� UTR�^(q��)��Ҳ�д�����m6A���RNA��������c�S���D(zhu��n)䛺��{(di��o)�أ�����mRNAǰ�w���У�mRNA����ͷ��g�����еġ�reader��øYTH������ʮ����Ҫ�����á�RNA�����D(zhu��n)��ø��methyltransferase���܉��������N6λ�l(f��)������������METTL3��METTL14��WTAP��Vir�ȡ���ȥ����ø����FTO��ALKBH5��
��֪��С����̥�ɼ�(x��)���Ќ�METTL3�ó���knockout�����܉���ֹ��l(f��)���ֻ���MELLT3������ȱʧ��(du��)С����̥�ɼ�(x��)���İl(f��)�������������á�ͬ���ڹ�ω�У�METTL3��ͬԴ����Ime4���ó���Ҳ���Ё��������ã�����NOTCH��̖(h��o)ͨ·�ܓp�Ķ�Ӱ푹�ω��������������ģʽֲ��M�Ͻ��У�METTL3ͬԴ����Ath-MTA���ó�����l(f��)��Ҳ��(hu��)�ܵ��ܴ�Ӱ푡����⣬Ime4����?q��)���ĸ�Ĝp��(sh��)����Ҳ�к���Ҫ�����á��N�N�E�������m6A���(du��)�������������ٰl(f��)���Լ����ڵ���̥�l(f��)�����P(gu��n)��Ҫ��������ĽY(ji��)��(g��u)����W(xu��)�о�������METTL3��METTL14��(hu��)�γ��s�j(lu��)����ʹ�����ã����������w��(n��i)�����ÙC(j��)��������Ȼδ֪�����cRNA������ø������
|
�(l��i)�e |
���� |
���� |
|
Writers |
METTL3, METTL14, WTAP, KIAA1492 |
METTL3��METTL14�γ��s�j(lu��)����w��(n��i)���w���RNA�l(f��)��m6A���������WTAP��Fl(2)d����KIAA1492��Vir���Լ�����factorsҲ�s�j(lu��)�����Ҫ�M�ɲ��֡��@ЩWriters����consensus motif |
|
Erasers |
FTO, ALKBH5�Լ�����homologies |
ȥ����ø���錧(d��o)m6Aȥ������� |
|
Readers |
YTHDF1��YTHDF2��YTHDF3�Լ�����ͬԴ���嵰�� |
�R(sh��)�eRNA���������Ϣ�������c����RNA�ķ��g��������^(gu��)�̡��@ЩYTHDF����ĵ����ձ鶼����YTH domain |
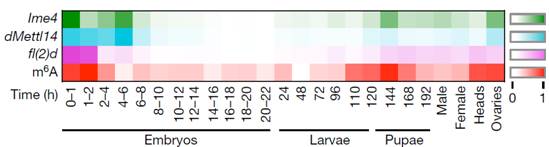
���x(ch��ng)����(g��)�������ڰ�����̥�ڡ����x(ch��ng)�ڣ�һ�g���x(ch��ng)�����g���x(ch��ng)����Ӽ���Լ����x(ch��ng)�ڡ������șz�y(c��)��Ұ����ω����(g��)�r(sh��)�ڵ�m6A���w�ļ���ˮƽ��ʹ���|(zh��)�V����(du��)Ime4��dMettl14��fl(2)d���N��������(du��)���_(d��)ˮƽ����һ��(g��)��D���ğ�D���҂����l(f��)�F(xi��n)����̥����������2С�r(sh��)��m6Aˮƽ̎��һ��(g��)�ܸߵ�ˮƽ��֮���_(k��i)ʼ�½��������g���x(ch��ng)�ڣ�m6Aˮƽ�_(k��i)ʼ��������Ӽ�����_(d��)����һ��(g��)�߷����u�½�����ω���x(ch��ng)�ڵ��^���Լ��ѳ���m6AˮƽҲһֱ�S������(du��)�^��ˮƽ��
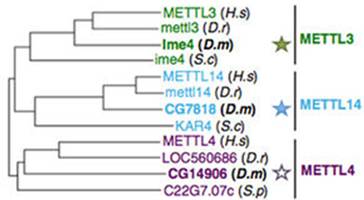
ϵ�y(t��ng)�M(j��n)����(sh��)������������ω��METTL3ͬԴ����Ime4�ЃɷN�������P(gu��n)�ĵ���CG7818��CG14906���ڹ�ω��SR2+��(x��)����embryonic-derived Schneider�����õ�Ime4��CG7818��m6Aˮƽ�½�70%���õ�CG14906�s�](m��i)���κ�Ӱ푡�����CG7818�cMETTL14�Y(ji��)��(g��u)�Ͼ��кܴ�ı����Ժ������ԣ����ߌ�CG7818����������dMettl14��ͬ��fl(2)d��Vir�����cWTAP��KIAA1429�߶�ͬԴ���õͺ�(du��)��ω��m6AˮƽҲ��(hu��)�a(ch��n)���^���Ӱ푡�
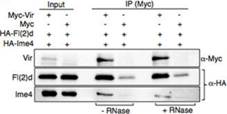
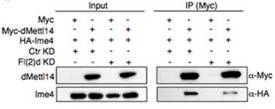
�ڲ����(d��ng)�����������w��(n��i)�У�RNA�����D(zhu��n)��øMETTL3��METTL14��(hu��)�γ��s�j(lu��)���WTAP��KIAA1429Ҳ����Ҫ�ĽM�ɲ��֡��ڹ�ω�����D(zhu��n)��ø��ͬ�r(sh��)�l(f��)�F(xi��n)��Vir��fl(2)d��Ime4�����ߌ�(du��)��ω��fl(2)d�õͺ�Ime4��dMettl14��interaction�@�����͡�
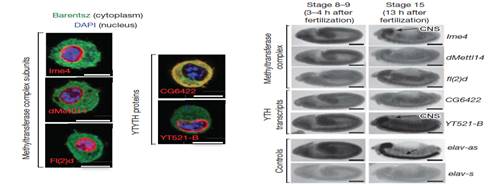
����λ��(sh��)�(y��n)�������cm6A������ø��λ�ڼ�(x��)���˃�(n��i)�Ҷ�����̥���ڰl(f��)���A��̎�ڸ߱��_(d��)ˮƽ��������̥���ڣ��@Щø����(j��ng)���ߌ�Ҳ���^�ߵ��D(zhu��n)�ˮƽ��
3.YT521-B�錧(d��o)m6A�{(di��o)��mRNA��׃����
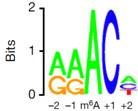
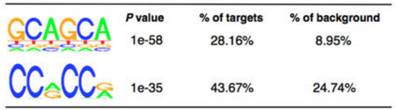
����ʹ��MeRIP-seq����812��(g��)�������b���˵���1120��(g��)m6A��peak������92%��peak������RRACH motif�����⣬�����(l��i)�͵����л�motifsҲ�����^�ߵ�m6Aˮƽ���@�����@Щmotifs����Ҳ��(du��)�����D(zhu��n)��ø�����خ��ԡ�
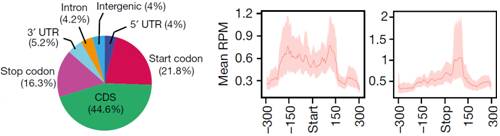
���⣬�c������������(l��i)�ƣ��ڹ�ω�w��(n��i)start codon��stop codon�^(q��)����ںܸߵ�m6A�������
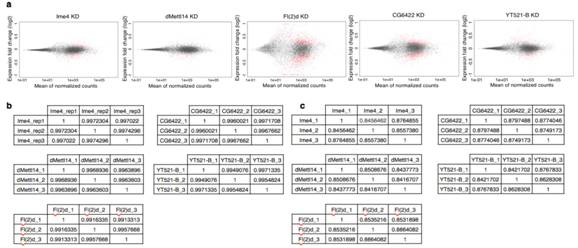
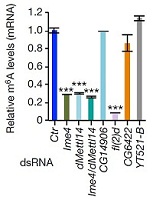
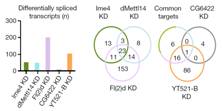
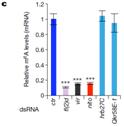
����(l��i)������ᘌ�(du��)SR2+��(x��)��ϵ���cm6A���P(gu��n)�ĸ��Nø�õͺ��M(j��n)�����D(zhu��n)䛽M�y(c��)���@Щø����Ime4��CG6422��dMellt14��YT521-B�Լ�Fl(2)d�����ψDa���҂��������@������MA-plot�@ʾ��ͬ�õ͗l���£�SR2+��(x��)��ϵ�еĻ�����_(d��)�l(f��)�����@���IJ��p<0.05�����Db��ʾ��ͬ�ó��l��������(g��)����W(xu��)�؏�(f��)������_(d��)��spearman���P(gu��n)ϵ��(sh��)�����Dc�t�Ǽ���ģʽ��spearman���P(gu��n)ϵ��(sh��)����
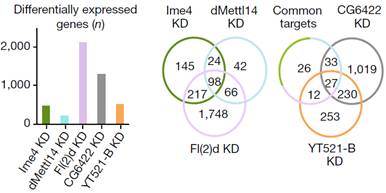
�������߰l(f��)�F(xi��n)Fl(2)d�õͺ�Ч��������@�����^(gu��)2000��(g��)������_(d��)�@������õ�Ime4��dMettl14��ͨ�^(gu��)������f���D���l(f��)�F(xi��n)SR2+�в���_(d��)����(sh��)��С��Fl(2)d��milder effects����
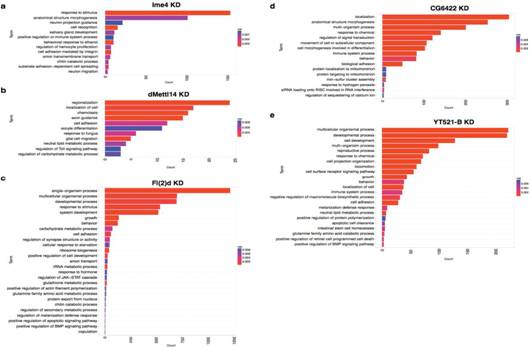
GO���ܸ��������������@Щ����_(d��)�Ļ����漰���N��������x���D(zhu��n)�\(y��n)����(x��)��ճ���ȡ��M��SR2+��(x��)��ϵ������ֱ����Դ����(j��ng)ϵ�y(t��ng)�������S������������neuronal functions�ϣ���axon guidance��synapse activity��
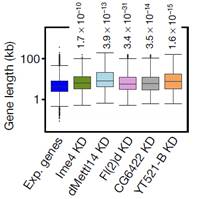
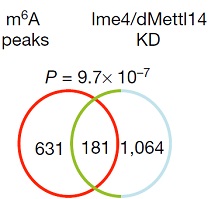
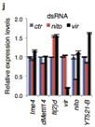
ͨ�^(gu��)���_(d��)�����ΈD�l(f��)�F(xi��n)�������^�ͽM�ļ�(x��)�������w���_(d��)ˮƽ�@������control�M������(l��i)����(du��)SR2+��(x��)��ϵ�е�Ime4��dMettl14�@2��(g��)�����M(j��n)���p�õͣ�double knockdown�����f���D�@ʾ����15%�Ļ�����m6A��peak��
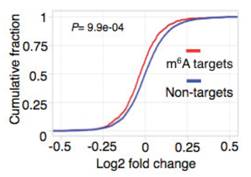
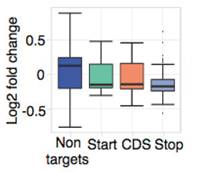
Ime4��dMettl14�p�ú�m6A����ˮƽ��Ȼ�l(f��)�����p������Ȼ�@���IJ���@��(g��)�Y(ji��)������ÿ��(g��)�D(zhu��n)䛱����ܓ��Ъ�(d��)����m6A peakλ�c(di��n)��
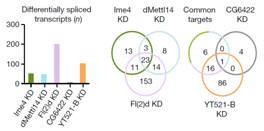
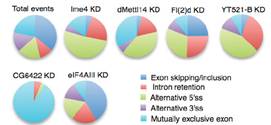
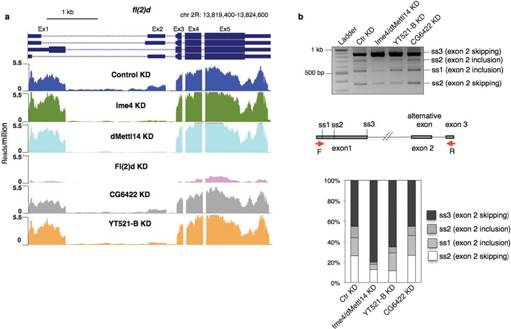
����߀�l(f��)�F(xi��n)�ڻ�������¼��ϣ��p�õĽY(ji��)���c2�Nø�Ϊ�(d��)�õĽY(ji��)����߀�Ǵ����@����ġ�������һ�Nø���õ͌�(sh��)�(y��n)�У�fl(2)d�@��(g��)������ʹʮ����Ҫ�����á�ͨ����r�£���ͬø�õͶ���(hu��)��(du��)����5���ļ����Լ���(n��i)���ӱ������О�a(ch��n)��Ӱ푣��@�N��rͬ��Ҳ�������˼�(x��)��ϵ�С�
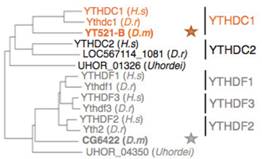
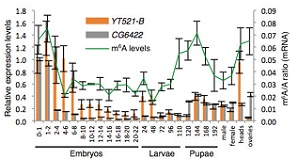
YTH���嵰����һ�Nʮ����Ҫ��readerø����ω�У�CG6422��YT521-B����YTH�����ͬԴ���ס�����(x��)����λ��(sh��)�(y��n)����CG6422λ�ڼ�(x��)���|(zh��)�У��ܾ�������̥��2С�r(sh��)�_(d��)�����ֵ������(l��i)���m(x��)�½����������x(ch��ng)�ں�Ӽ��һֱ�S���^�͵�ˮƽ��Ȼ��������(x��)����λ��(sh��)�(y��n)����YT521-Bλ�ڼ�(x��)�����У���������̥����(j��ng)ϵ�y(t��ng)�ѽ�(j��ng)���x(ch��ng)�Ĵ��X��̎���^�ߵ��D(zhu��n)�ˮƽ��
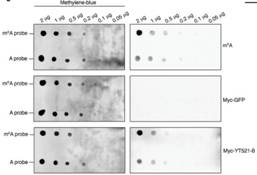
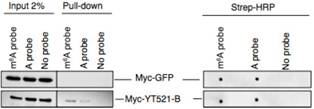
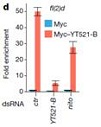
ʹ���c(di��n)ӡӛ����dot-plot����pull down��(sh��)�(y��n)������YT521-B�܉��cRNA�ϵ�m6Aλ�c(di��n)��Y(ji��)�ϡ��D(zhu��n)䛽M�y(c��)��������õ�CG6422�����RNA�����¼���Ӱ푲������õ�YT521-B���D(zhu��n)䛽M�Y(ji��)��������103��(g��)����ļ����¼���@�����f���D�@ʾYT521-B�ccommon target�Լ�CG6422��������30%�������@Щ�C��(j��)������YT521-B��pre-mRNA��������ʹ�O����Ҫ�Ĺ��ܡ�
4.m6A���D(zhu��n)��ø��(du��)��ω���О��{(di��o)��
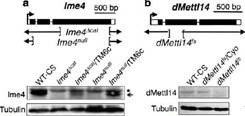
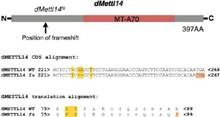
���ˌ���m6A���P(gu��n)ø�ĝ��ڹ��ܣ����߷քe��(g��u)����Ime4��dMellt14�ó��Ĺ�ω��
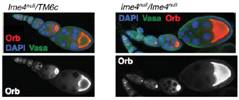
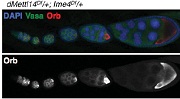
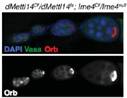
����ᘌ�(du��)Ime4�@��(g��)����ʹ�ÃɷN�����ó�������Ime4Null��CDSȫ���ó�����Ime4Cat��C�˱������Y(ji��)��(g��u)�����°l(f��)�F(xi��n)�o(w��)Փ�Ǽ��Ϲ�ω߀�ǰl(f��)���s��ͻ׃�Ĺ�ω�����x(ch��ng)��������������ڡ��ѳ��Л](m��i)�аl(f��)�F(xi��n)�κ�encapsulation defects��

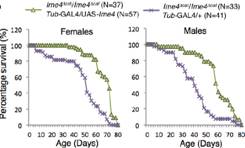
����ͻ׃��ω����׃�̣��w�к��������裬�o(w��)���ϳᡣ�M��ͻ׃��ω�܉��㏊(qi��ng)�ϳ���s����Ǻ��y�����ϔn�ڸ������ز��ı��档����dMettl14����ͻ׃��ω�mȻ��l(f��)�������������О������s�ѽ�(j��ng)ȱʧ�������(y��n)�CIme4��dMettl14�Ƿ��ڹ�ω�w��(n��i)�܉���a(b��)�������ߘ�(g��u)�����pͻ׃��ω�͆λ���ͻ׃��ω���Y(ji��)���������@2��(g��)�����������{(di��o)�ص�ͨ·Ҳ���ƣ������{(di��o)��common targets�����⣬Ime4�ƺ��ڹ����ϱ�dMettl14��ռ��(y��u)��(sh��)���ִ�����(y��ng)��Ҫ�����D(zhu��n)��ø���ض��Ĵ��Y(ji��)��(g��u)��
С�Nʿ����ω�������x(ch��ng)�V�p��Ŀ��ֻ��һ��(du��)�����һ��(du��)����ѽ�(j��ng)�M(j��n)����ƽ����������o��ƽ���w�С�ͨ������(hu��)�ϔn���ز������ı��档
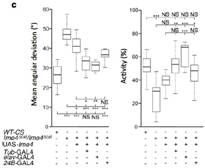

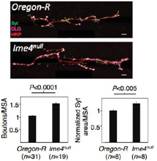
�О錍(sh��)�(y��n)����Ime4ͻ׃��ω�����ٶ��@������Ұ����ω������������Orientation��Ҳ���F(xi��n)��(w��n)�}���N�N�C��(j��)����m6A���ͨ�^(gu��)�{(di��o)����(j��ng)ϵ�y(t��ng)���Ķ���(y��n)��Ӱ푹�ω���О顣����ͨ�^(gu��)�����О�̽����ӵ���(j��ng)ȱ�ݺͷ��әC(j��)�ƣ�������Ime4ͻ׃��ω���x(ch��ng)���M(j��n)���˼�����(j��ng)���^�y(c��)ԇ��NMJ�����ψD�@ʾNMJ��Ime4ͻ׃�����x(ch��ng)��(n��i)��(sh��)ͻ���L(zh��ng)��Ұ���͵�1.5�����Y(ji��)������Ime4ͨ�^(gu��)����NMJ�Ę�(sh��)ͻ���L(zh��ng)��(l��i)�{(di��o)��(ji��)��ω���x(ch��ng)�������О顣
�����b����ω�О�İл������ߌ�(du��)�����g�ƹ�ω���x(ch��ng)���^���M(j��n)���˽��ʲ��M(j��n)���D(zhu��n)䛽M�y(c��)��ͨ�^(gu��)�D(zhu��n)䛽M�Y(ji��)��������������Ime4ͻ׃�����b����1600����(g��)������ڻ������ģʽ�ϰl(f��)����׃���@Щ�����^�ֶ��ǿ��ƹ�ω�О�ġ�����(l��i)������(du��)SR2+��(x��)��ϵԭ�е�MeRIP��(sh��)��(j��)�c�D(zhu��n)䛽M��(sh��)��(j��)�M(j��n)�бȌ�(du��)���l(f��)�F(xi��n)���{(di��o)�ع�ω�О�Ļ��l(f��)����m6A������������Ɯy(c��)�������D(zhu��n)��øȱ�ݵ���r�£������в�ֹһ��(g��)�����{(di��o)�ع�ω���О顣
5.�������m6A�{(di��o)�ع�ω�Ԅe�ֻ�
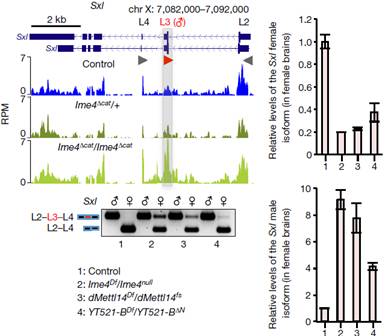
�ڹ�ω�w��(n��i),sxl�����@��(g��)�_(k��i)�P(gu��n)�����c�������εĄ����a(b��)��Ч��(y��ng)��(l��i)�{(di��o)�ؿ��ƹ�ω���Ԅe�ֻ�����ω�Ԅe�ֻ��C(j��)��ԔҊ(ji��n)����С�Nʿ���֡�����(n��i)������ͨ�^(gu��)��(sh��)�(y��n)�l(f��)�F(xi��n)Ime4��(du��)SxlӰ푺ܴ�Sxl���������x(ch��ng)�ʹ��x(ch��ng)��ͬ�r(sh��)���ڣ�ֻ���^(gu��)���x(ch��ng)Sxl������һ��(g��)�~���������@�ӡ���ͻ׃�w���Թ�ω��������Ime4��Ӱ푣����ɷNͻ׃�ʹ��Թ�ω�ڻ������϶����F(xi��n)���c���Թ�ω���е����@�Ӳ����Լ����۹�ω��(d��)�еļ���ģʽ���o(w��)Փ�Ǵ��x(ch��ng)߀�����x(ch��ng)��Sxl��ͬ�D(zhu��n)䛱��½�ˮƽ�����e���@���@����m6A�����ڴ��X���خ��Ը�����
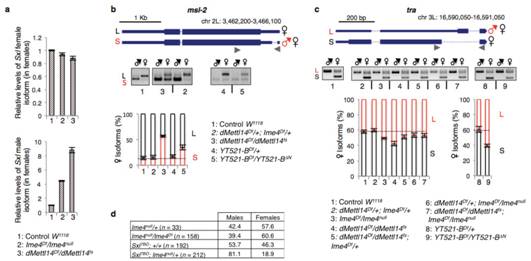
Sxl���������{(di��o)�ص�2��(g��)����tra��mslҲ�l(f��)�����@��׃�����@���������D(zhu��n)��ø���ܕ�(hu��)Ӱ�Sxl�����pre-mRNA�ļ��У��Ķ�Ӱ푹�ω�Ԅe�ֻ��̈́����a(b��)��Ч��(y��ng)�������(y��n)�C�@��(g��)���룬����ʹ��Ime4�s��ͻ׃�Ĵ��x(ch��ng)�cSxl�s��ͻ׃�����x(ch��ng)�M(j��n)���s�����y(t��ng)Ӌ(j��)�������ʰl(f��)�F(xi��n)������x(ch��ng)����Ӱ푣�Ime4�cSxlȱʧ�Ĵ��Դ�����½������ԽY(ji��)��������Ime4�Ɖ��˹�ω�w��(n��i)�����Ą����a(b��)��Ч��(y��ng)����ͨ�^(gu��)�{(di��o)��SxlӰ푴��۹�ω�Ĵ�
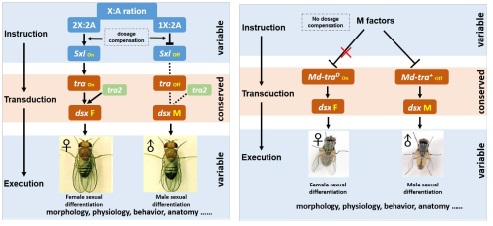
С�Nʿ����ω(��)�ͼ�ω(��)�ڿ����Ԅe�ֻ��ϲ�����ȫ��ͬ�ęC(j��)�ơ�2017��Science����Փ���״��b���˼�ω���ԛQ������Mdmd����ωͨ�������a(b��)�������C(j��)�ƣ�dosage compensation��������(d��ng)���ε�X:A�ij�Ⱦɫ�w����Ⱦɫ�w�����Q�����ε�Sxl�Ƿ��(hu��)��������Sxl�����ܵ�Ӱ푣��o(w��)Փ���ε�X:AȾɫ�w������Σ���ω����(hu��)�l(f��)�������Գ��x(ch��ng)���෴�����Sxlһֱ̎�ڼ����B(t��i)���t��ω��(hu��)�l(f��)���ɴ��Գ��x(ch��ng)��
6.YT521-Bȱʧ�cm6A����֮�g���P(gu��n)ϵ
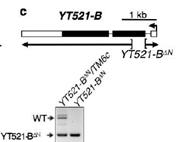
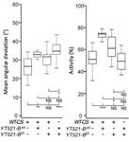
�b��YT521-B�܉��خ����R(sh��)�em6A����λ�c(di��n)���Ķ�Ӱ�SR2+��(x��)���н^����m6A�錧(d��o)��RNA�����¼������ߌ�(du��)���N���cRNA������ø�M(j��n)�����ó���knockout����(sh��)�(y��n)��YT521-Bȱʧ�܉�����ͬ�D(zhu��n)䛱����_(d��)��������YT521-Bͻ׃��ω�mȻ�܉��������x(ch��ng)�ڣ������w�е��О��ܵ��˺ܴ��Ӱ푡�
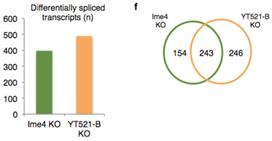
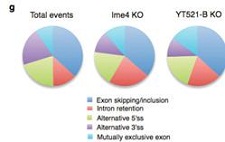
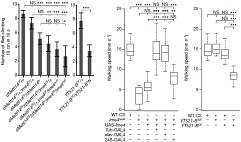
��YT521-Bͻ׃���x(ch��ng)�cIme4ͻ׃���x(ch��ng)�M(j��n)���D(zhu��n)䛽M�y(c��)�l(f��)�F(xi��n)������243��397��(g��)������ڼ���ģʽ�IJ��5���˿�׃����λ�c(di��n)�خ��Բ��ߣ���(n��i)���Ӳ��ֱ�����Ȼ�^�ࡣ�@Ȼ��YT521-Bͻ׃Ҳ��Ӱ�male-specific Sxl���Ķ�Ӱ����ε�tra��msl-2���^��ʹ�ô��x(ch��ng)�w��(n��i)female-specific Sxl�D(zhu��n)�ˮƽ���͡����ϽY(ji��)������m6A���D(zhu��n)��ø��ͨ�^(gu��)YT521-B�{(di��o)�ع�ω�О���Ԅe�ֻ���
7.Nito��һ�Nȫ�µą��cm6A���ø
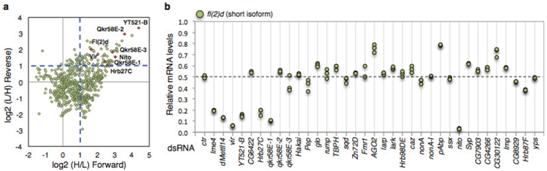
����̽��YT521-B���{(di��o)�ؙC(j��)�ƣ���������SILAC���g(sh��)����(du��)SR2+��(x��)�����B(y��ng)�^(gu��)���еİ������M(j��n)��ͬλ�ؘ�(bi��o)ӛ���Y(ji��)���|(zh��)�V���g(sh��)�����߹����팦(du��)SR2+��(x��)���к�YT521-B���߽Y(ji��)�ϵĵ����M(j��n)�ж�����������һ�루n=30���c�A(y��)�y(c��)�ĽY(ji��)�ϵ�������ϡ������(y��n)�C�����Ƿ����m6A�錧(d��o)�ļ����¼����҂��քe��(du��)�@30��(g��)�����M(j��n)����depleted��Ȼ���u(p��ng)��fl(2)d��(du��)�����¼���Ӱ푡����е���Hrb27C��Qkr58E-1��Nito�cfl(2)d���Ƶļ��ЬF(xi��n)������ϡ�
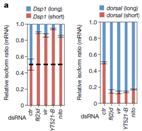
������6��(g��)�~���m6A�錧(d��o)�ļ����¼�������Hrb27C��Qkr58E-1�H�H�{(di��o)��һ��(g��)���ׁ�������Nito��ȱʧ��(hu��)��(d��o)�»�����Еr(sh��)�g�������@��(g��)�����F(xi��n)���cYT521-B���䵰��(f��)���﹦��ȱʧ��ĬF(xi��n)����^���ơ�
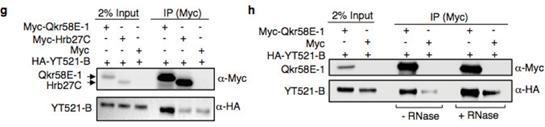
����(l��i)����ʹ�����߹����팢YT521-B�cHrb27C��Qkr58E-1��Nito���N�Y(ji��)�ϵ����M(j��n)�й�����̽�������g������á��Y(ji��)��������Hrb27C��Qkr58E-1�cYT521-B�ĽY(ji��)����ه(l��i)��RNA��
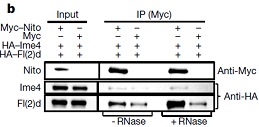
��Nito�����cYT521-B������Y(ji��)�τt��ȫ����ه(l��i)��RNA������Nito�����cIme4�Լ�fl(2)d��Y(ji��)����ه(l��i)��RNA��Nito������õ͕�(hu��)��(d��o)��m6A���ˮƽ�Ľ��͡����Nitoȱʧ��(du��)��m6A��ˮƽӰ푺ܴ�ԓ�о�߀�b�����˹�ωm6A���D(zhu��n)��ø��(f��)���w�е�����Nito(Spenito)��
����ͨ�Ź���̖(h��o)
���Մ�(d��ng)�B(t��i) | �˲��Ј�(ch��ng) | �¼��g(sh��)��(zhu��n)�� | �Ї�(gu��)�ƌW(xu��)�� | ��չ�_(t��i) | BioHot | ���v��ֱ�� | ��(hu��)չ���� | �r(ji��)��(zhu��n)�� | ���g(sh��)��Ӎ | ���M(f��i)ԇ��
���(qu��n)���� ����ͨ
Copyright© eBiotrade.com, All Rights Reserved
(li��n)ϵ���䣺
��ICP��09063491̖(h��o)
